

Identifying the core genome of the nucleus-forming bacteriophage family and characterization of Erwinia phage RAY
- PMID: 37120812
- PMCID: PMC10299810
- DOI: 10.1016/j.celrep.2023.112432
Identifying the core genome of the nucleus-forming bacteriophage family and characterization of Erwinia phage RAY
Abstract
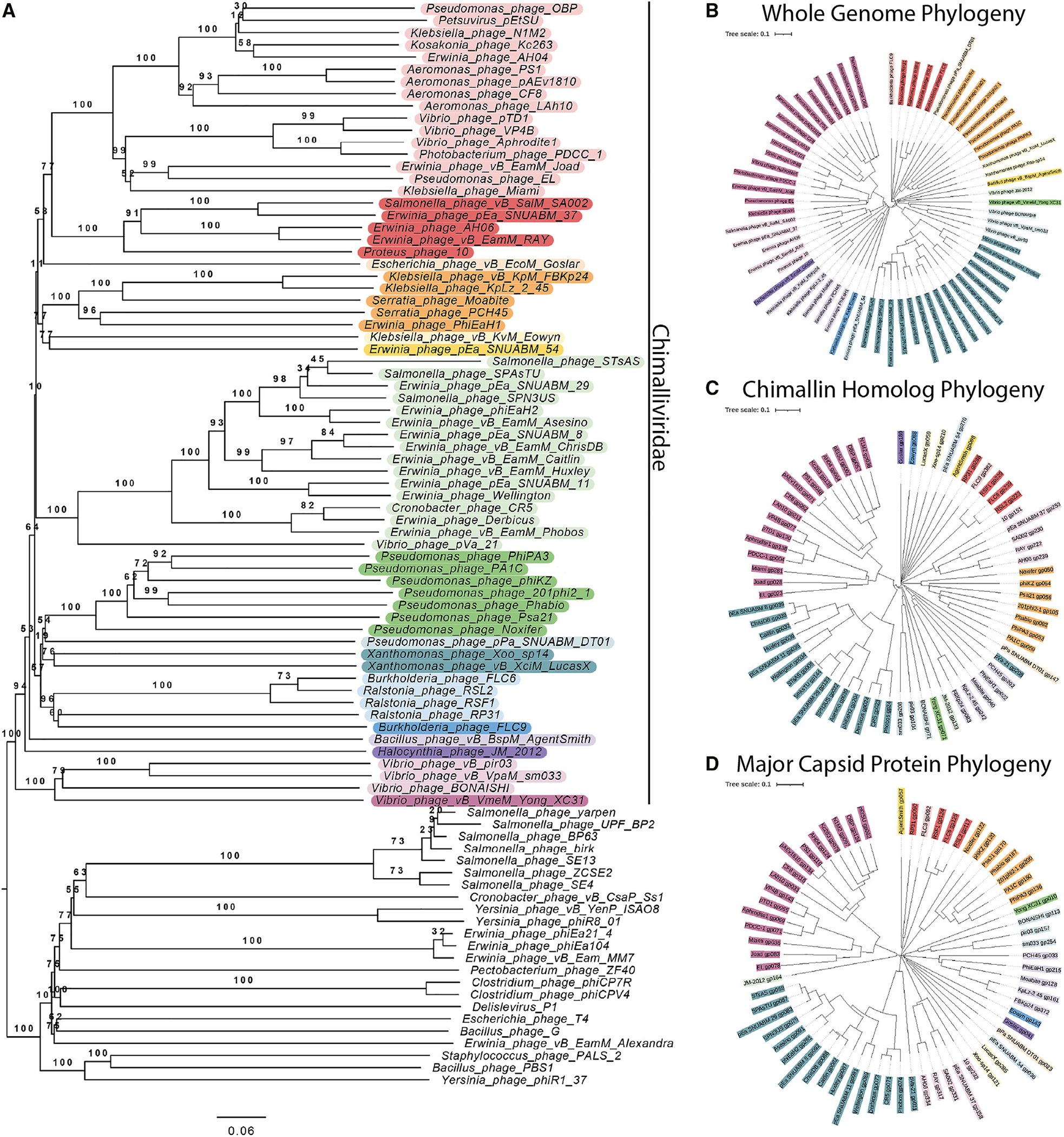
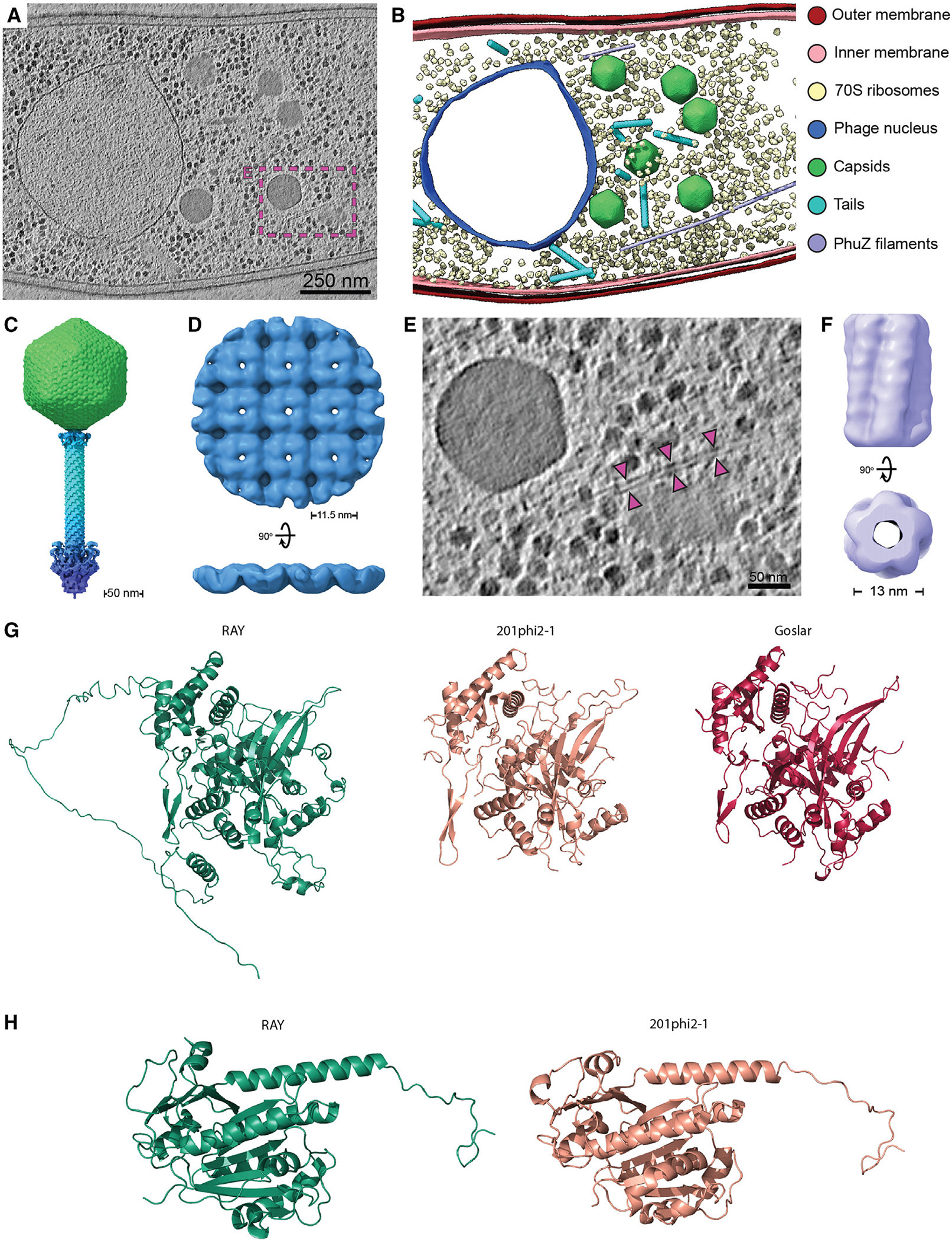
We recently discovered that some bacteriophages establish a nucleus-like replication compartment (phage nucleus), but the core genes that define nucleus-based phage replication and their phylogenetic distribution were still to be determined. Here, we show that phages encoding the major phage nucleus protein chimallin share 72 conserved genes encoded within seven gene blocks. Of these, 21 core genes are unique to nucleus-forming phage, and all but one of these genes encode proteins of unknown function. We propose that these phages comprise a novel viral family we term Chimalliviridae. Fluorescence microscopy and cryoelectron tomography studies of Erwinia phage vB_EamM_RAY confirm that many of the key steps of nucleus-based replication are conserved among diverse chimalliviruses and reveal variations on this replication mechanism. This work expands our understanding of phage nucleus and PhuZ spindle diversity and function, providing a roadmap for identifying key mechanisms underlying nucleus-based phage replication.
Keywords: CP: Microbiology; Chimallin; Erwinia; PhuZ; core genome; cytoskeleton; nuclear shell; phage; phage nucleus; phage tubulin.
Copyright © 2023 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests K.P. and J.P. have an equity interest in Linnaeus Bioscience Incorporated and receive income. The terms of this arrangement have been reviewed and approved by the University of California, San Diego, in accordance with its conflict-of-interest policies.
Figures





Update of
-
Identifying the core genome of the nucleus-forming bacteriophage family and characterization of Erwinia phage RAY.bioRxiv [Preprint]. 2023 Feb 25:2023.02.24.529968. doi: 10.1101/2023.02.24.529968. bioRxiv. 2023. Update in: Cell Rep. 2023 May 30;42(5):112432. doi: 10.1016/j.celrep.2023.112432. PMID: 36865095 Free PMC article. Updated. Preprint.
References
-
- Labarde A, Jakutyte L, Billaudeau C, Fauler B, López-Sanz M, Ponien P, Jacquet E, Mielke T, Ayora S, Carballido-López R, and Tavares P (2021). Temporal compartmentalization of viral infection in bacterial cells. Proc. Natl. Acad. Sci. USA 118, e2018297118. 10.1073/pnas.2018297118. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
