

Ribosome-targeting antibiotics and resistance via ribosomal RNA methylation
- PMID: 37122541
- PMCID: PMC10131624
- DOI: 10.1039/d2md00459c
Ribosome-targeting antibiotics and resistance via ribosomal RNA methylation
Abstract
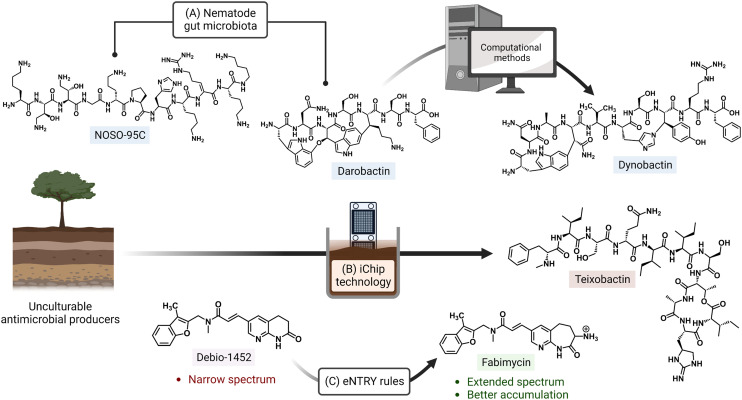
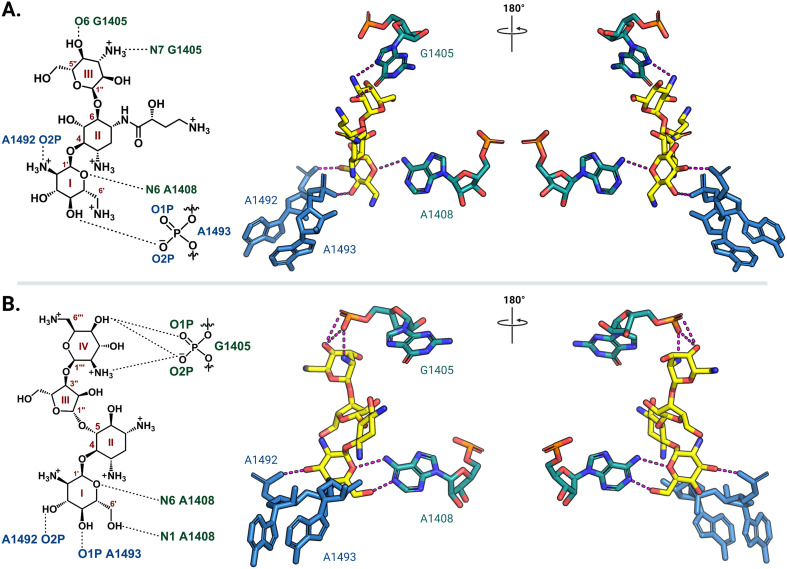
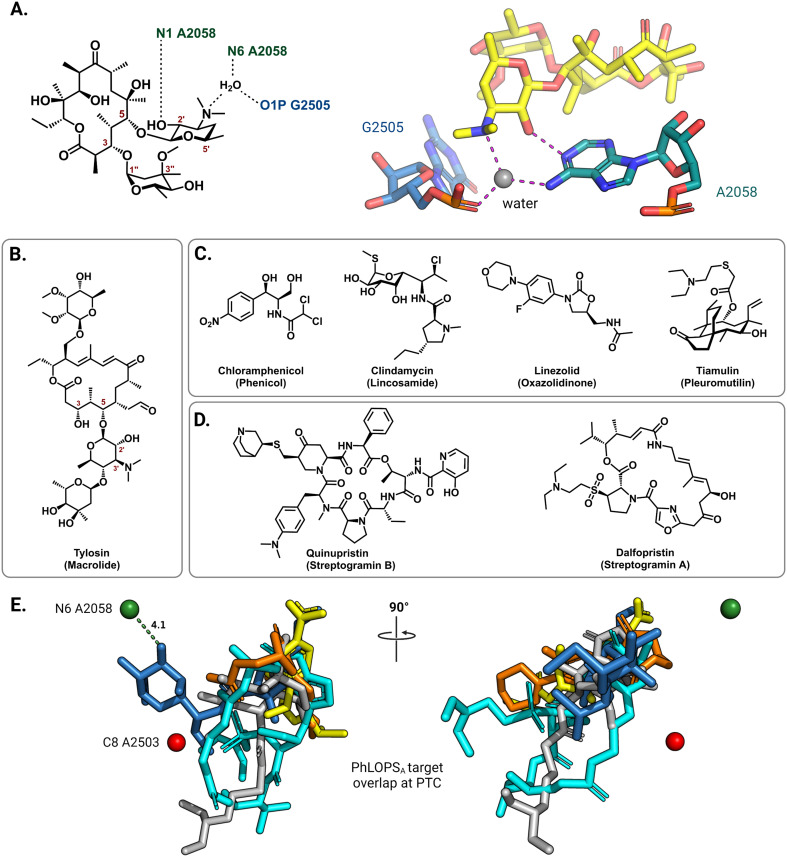
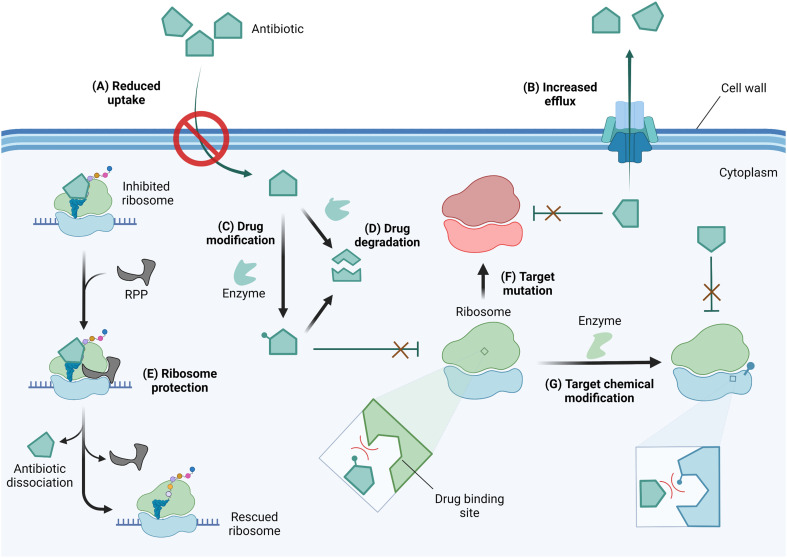
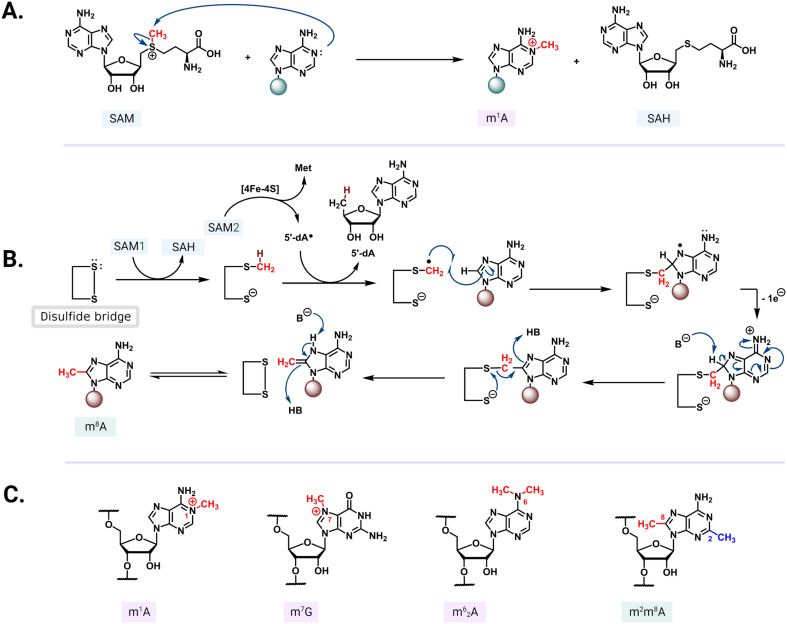
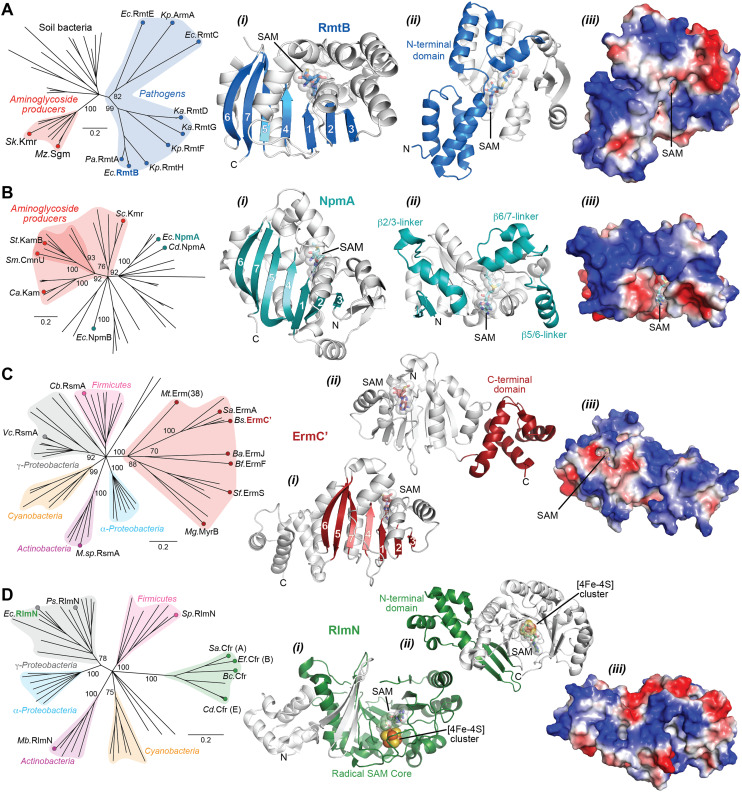
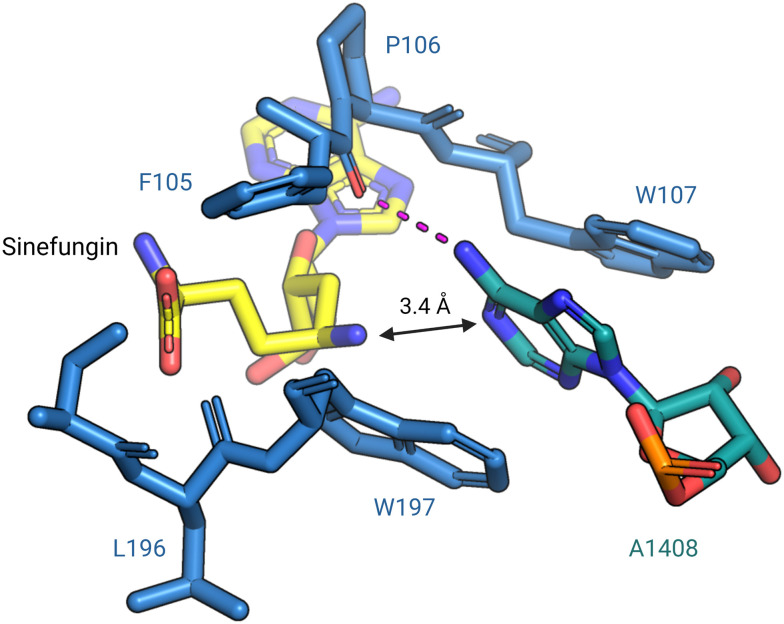
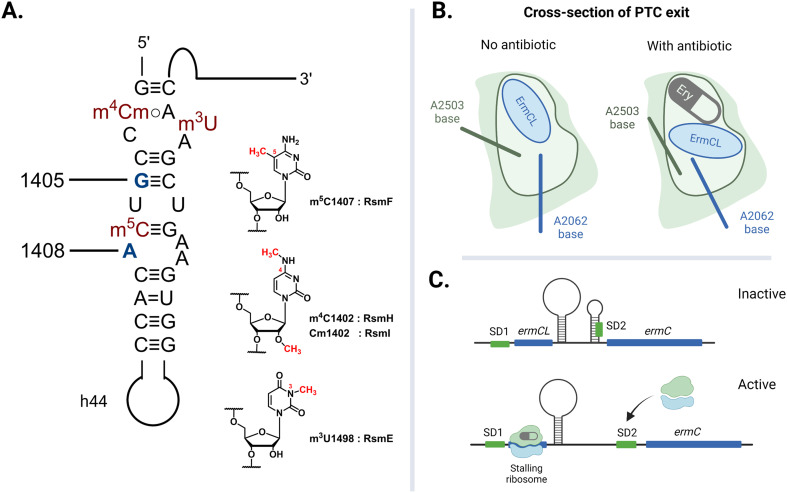
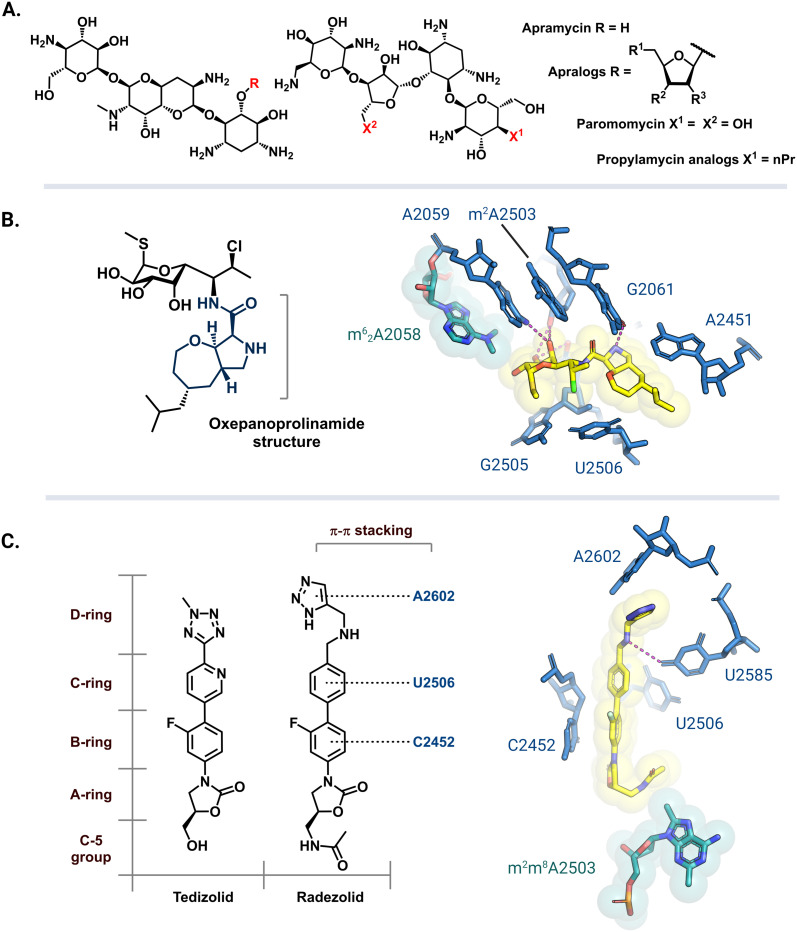
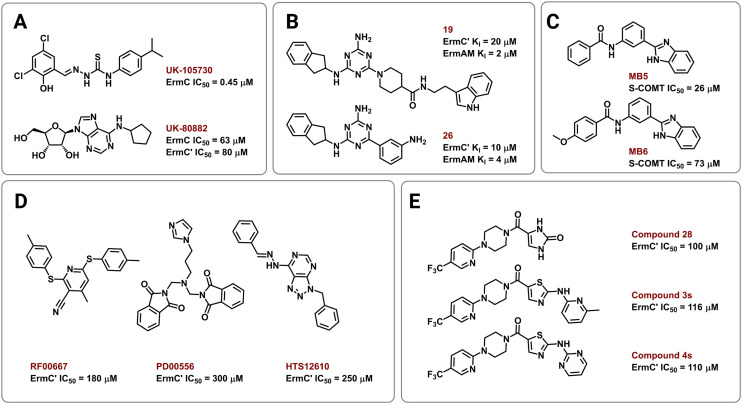
The rise of multidrug-resistant bacterial infections is a cause of global concern. There is an urgent need to both revitalize antibacterial agents that are ineffective due to resistance while concurrently developing new antibiotics with novel targets and mechanisms of action. Pathogen associated resistance-conferring ribosomal RNA (rRNA) methyltransferases are a growing threat that, as a group, collectively render a total of seven clinically-relevant ribosome-targeting antibiotic classes ineffective. Increasing frequency of identification and their growing prevalence relative to other resistance mechanisms suggests that these resistance determinants are rapidly spreading among human pathogens and could contribute significantly to the increased likelihood of a post-antibiotic era. Herein, with a view toward stimulating future studies to counter the effects of these rRNA methyltransferases, we summarize their prevalence, the fitness cost(s) to bacteria of their acquisition and expression, and current efforts toward targeting clinically relevant enzymes of this class.
This journal is © The Royal Society of Chemistry.
Conflict of interest statement
There are no conflicts to declare.
Figures
References
-
- Larsen J. Raisen C. L. Ba X. Sadgrove N. J. Padilla-González G. F. Simmonds M. S. J. Loncaric I. Kerschner H. Apfalter P. Hartl R. Deplano A. Vandendriessche S. Bolfíková B. Černá. Hulva P. Arendrup M. C. Hare R. K. Barnadas C. Stegger M. Sieber R. N. Skov R. L. Petersen A. Angen Ø. Rasmussen S. L. Espinosa-Gongora C. Aarestrup F. M. Lindholm L. J. Nykäsenoja S. M. Laurent F. Becker K. Walther B. Kehrenberg C. Cuny C. Layer F. Werner G. Witte W. Stamm I. Moroni P. Jørgensen H. J. de Lencastre H. Cercenado E. García-Garrote F. Börjesson S. Hæggman S. Perreten V. Teale C. J. Waller A. S. Pichon B. Curran M. D. Ellington M. J. Welch J. J. Peacock S. J. Seilly D. J. Morgan F. J. E. Parkhill J. Hadjirin N. F. Lindsay J. A. Holden M. T. G. Edwards G. F. Foster G. Paterson G. K. Didelot X. Holmes M. A. Harrison E. M. Larsen A. R. Nature. 2022;602:135–141. doi: 10.1038/s41586-021-04265-w. - DOI - PMC - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Chemical Information