

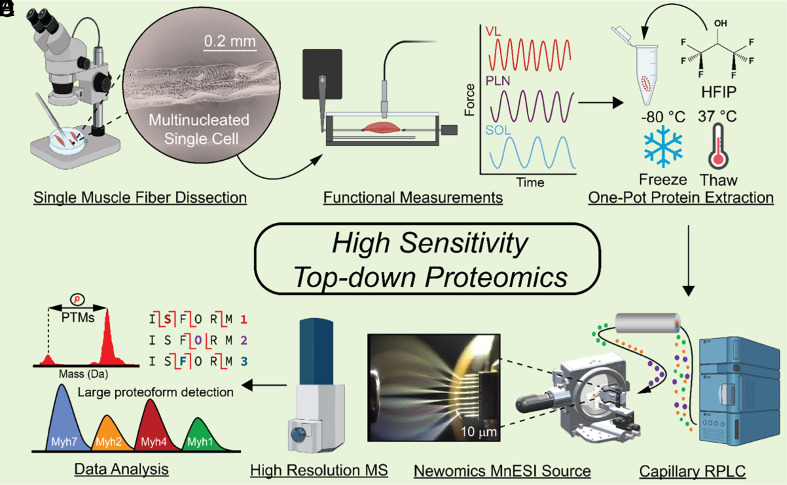
High sensitivity top-down proteomics captures single muscle cell heterogeneity in large proteoforms
- PMID: 37126723
- PMCID: PMC10175728
- DOI: 10.1073/pnas.2222081120
High sensitivity top-down proteomics captures single muscle cell heterogeneity in large proteoforms
Abstract
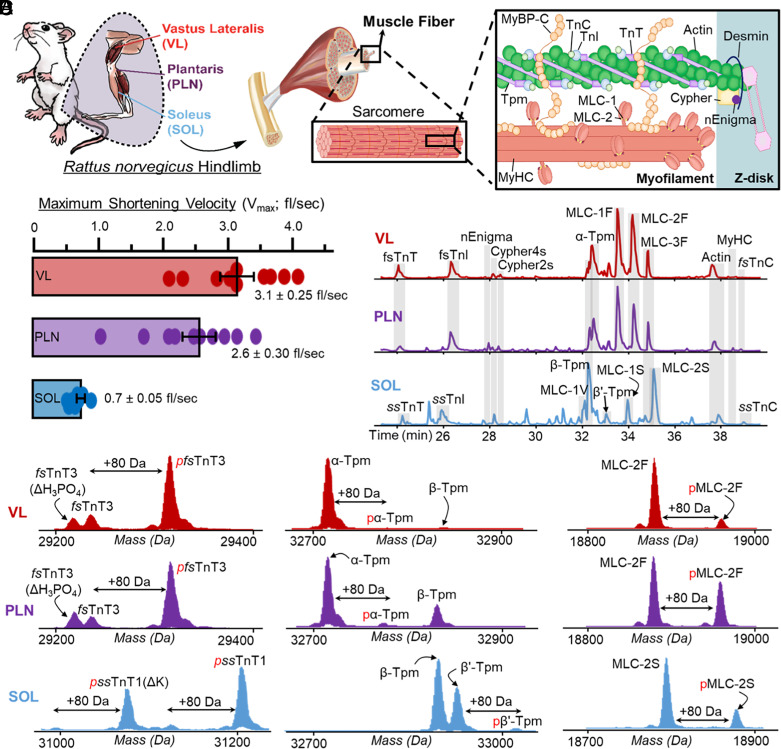
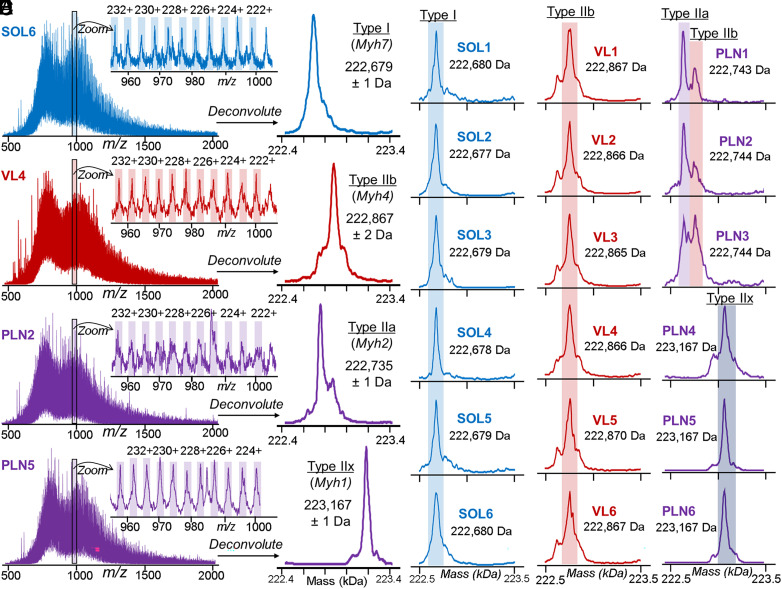
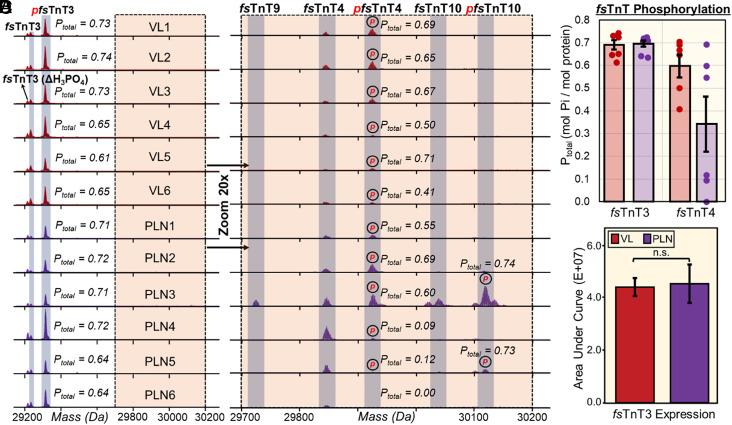
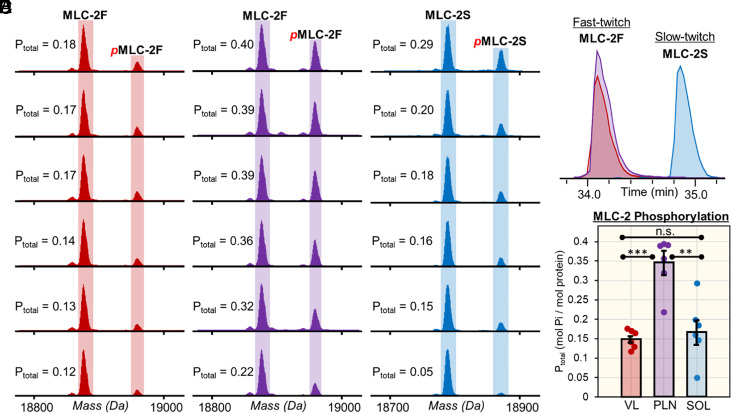
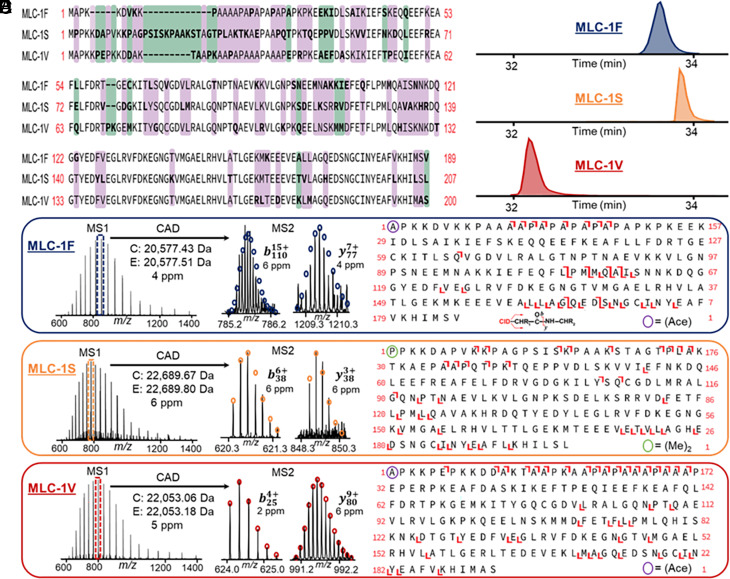
Single-cell proteomics has emerged as a powerful method to characterize cellular phenotypic heterogeneity and the cell-specific functional networks underlying biological processes. However, significant challenges remain in single-cell proteomics for the analysis of proteoforms arising from genetic mutations, alternative splicing, and post-translational modifications. Herein, we have developed a highly sensitive functionally integrated top-down proteomics method for the comprehensive analysis of proteoforms from single cells. We applied this method to single muscle fibers (SMFs) to resolve their heterogeneous functional and proteomic properties at the single-cell level. Notably, we have detected single-cell heterogeneity in large proteoforms (>200 kDa) from the SMFs. Using SMFs obtained from three functionally distinct muscles, we found fiber-to-fiber heterogeneity among the sarcomeric proteoforms which can be related to the functional heterogeneity. Importantly, we detected multiple isoforms of myosin heavy chain (~223 kDa), a motor protein that drives muscle contraction, with high reproducibility to enable the classification of individual fiber types. This study reveals single muscle cell heterogeneity in large proteoforms and establishes a direct relationship between sarcomeric proteoforms and muscle fiber types, highlighting the potential of top-down proteomics for uncovering the molecular underpinnings of cell-to-cell variation in complex systems.
Keywords: mass spectrometry; proteoform; proteomics; single cell; single muscle fiber.
Conflict of interest statement
A co-author, Dr. Daojing Wang, is an employee of Newomics, owns the stocks of Newomics, and is the founder and CEO of Newomics.
Figures
References
-
- Perkel J. M., Single-cell analysis enters the multiomics age. Nature 595, 614–616 (2021).
-
- R. Elmentaite, C. Domínguez Conde, L. Yang, S. A. Teichmann, Single-cell atlases: Shared and tissue-specific cell types across human organs. Nat. Rev. Genet. 23, 395–410 (2022). - PubMed
-
- Stuart T., Satija R., Integrative single-cell analysis. Nat. Rev. Genet. 205, 257–272 (2019). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
