

Targeting hepatic serine-arginine protein kinase 2 ameliorates alcohol-associated liver disease by alternative splicing control of lipogenesis
- PMID: 37129868
- PMCID: PMC10592686
- DOI: 10.1097/HEP.0000000000000433
Targeting hepatic serine-arginine protein kinase 2 ameliorates alcohol-associated liver disease by alternative splicing control of lipogenesis
Abstract
Background and aims: Lipid accumulation induced by alcohol consumption is not only an early pathophysiological response but also a prerequisite for the progression of alcohol-associated liver disease (ALD). Alternative splicing regulates gene expression and protein diversity; dysregulation of this process is implicated in human liver diseases. However, how the alternative splicing regulation of lipid metabolism contributes to the pathogenesis of ALD remains undefined.
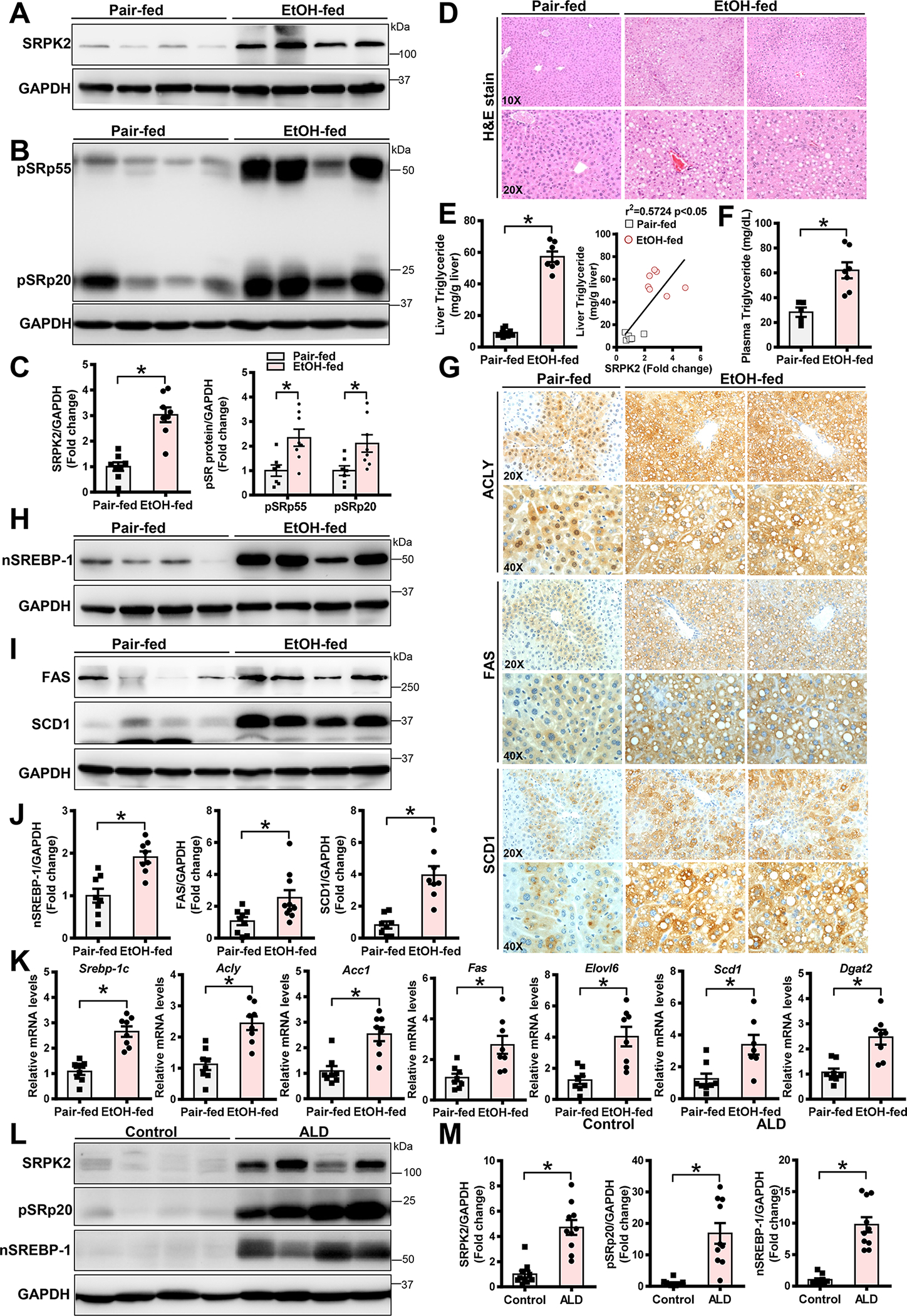
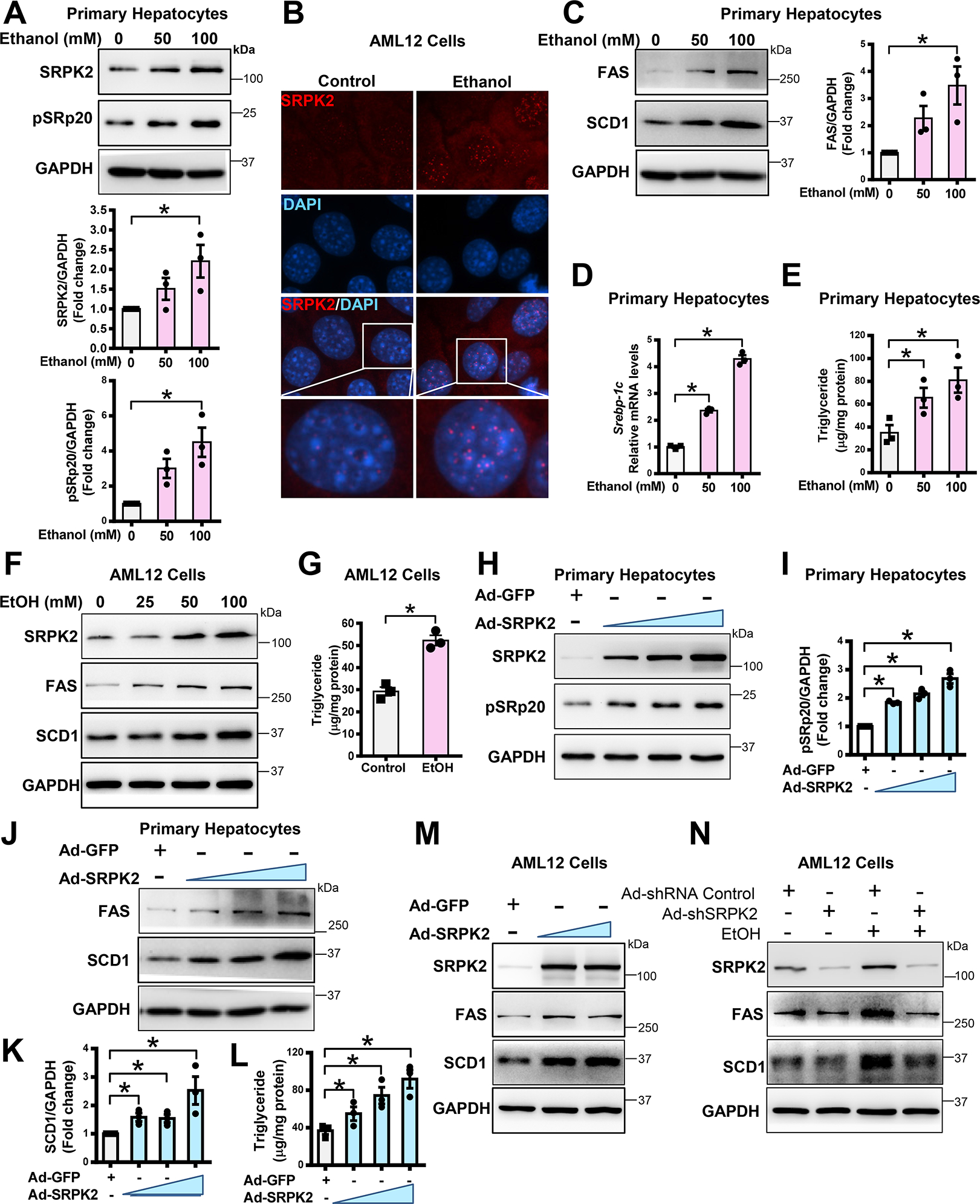
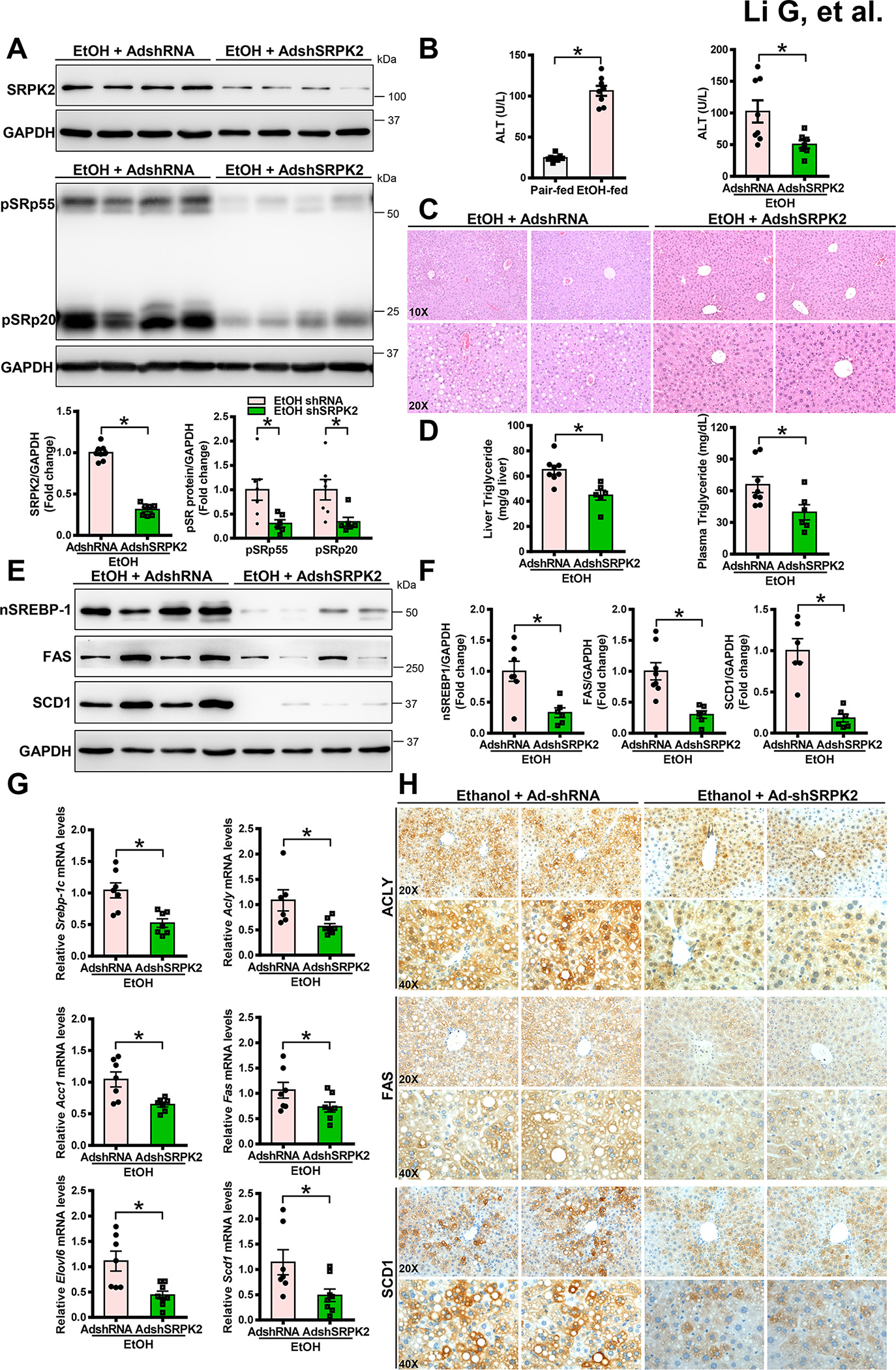
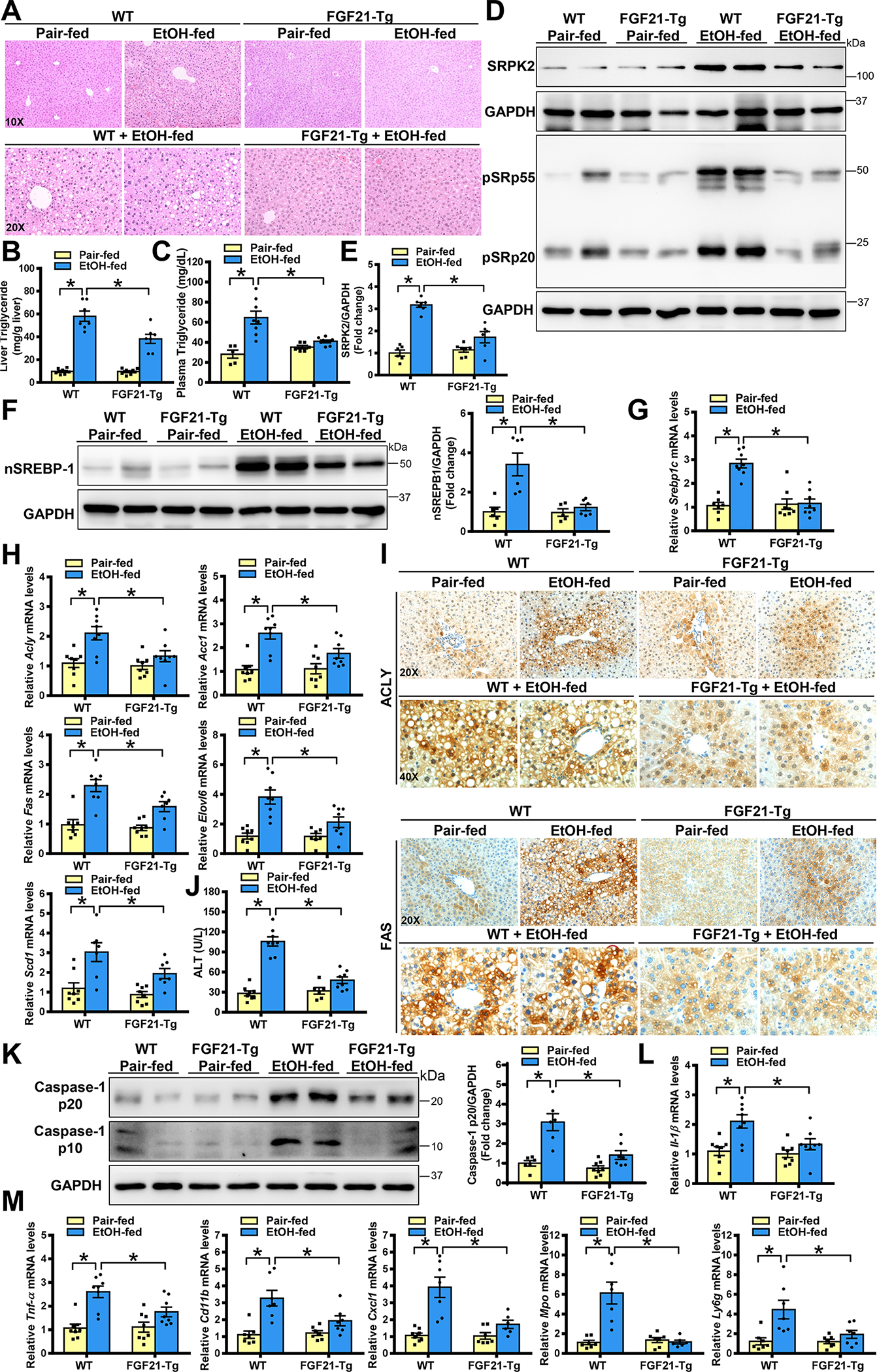
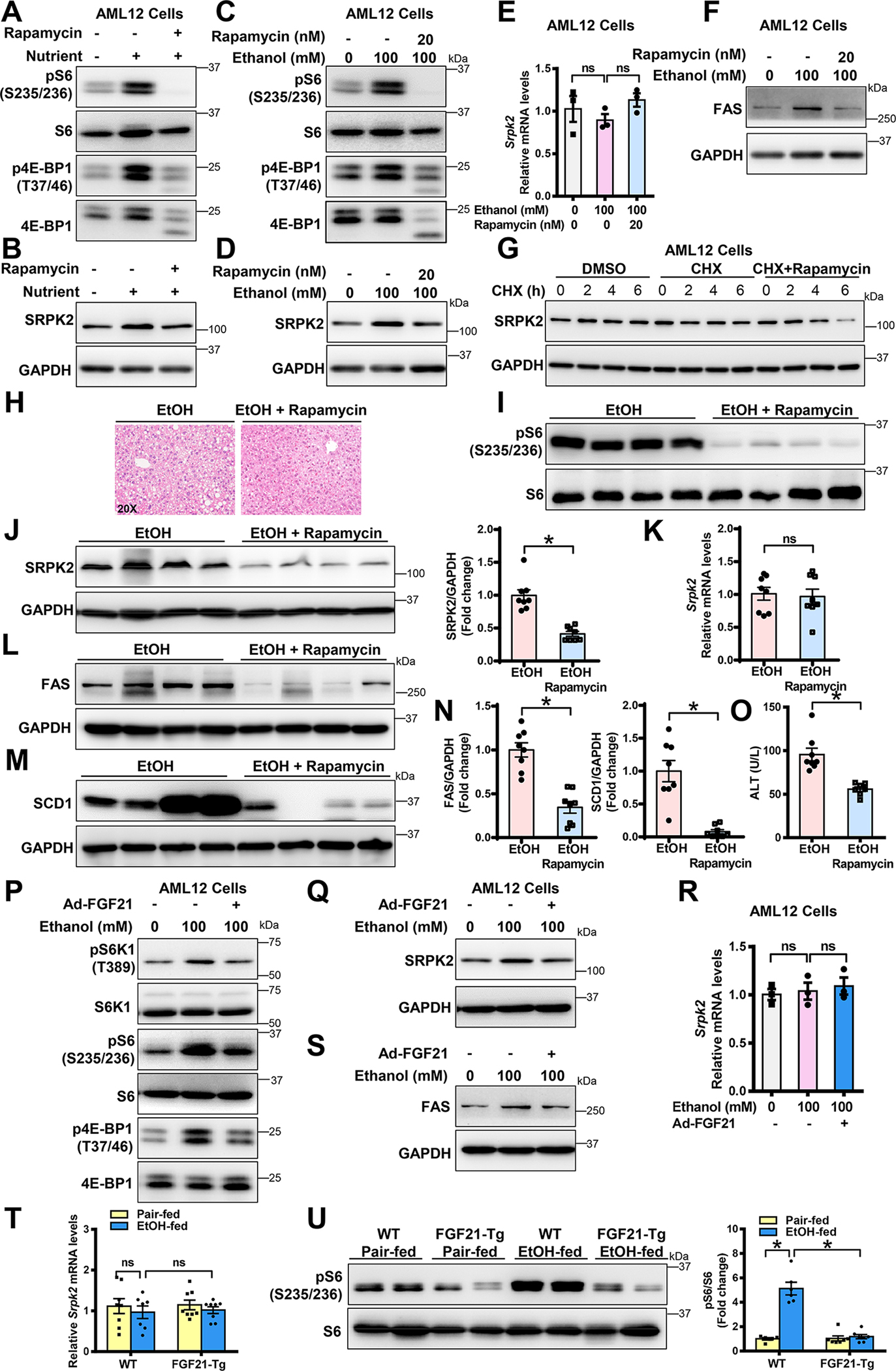
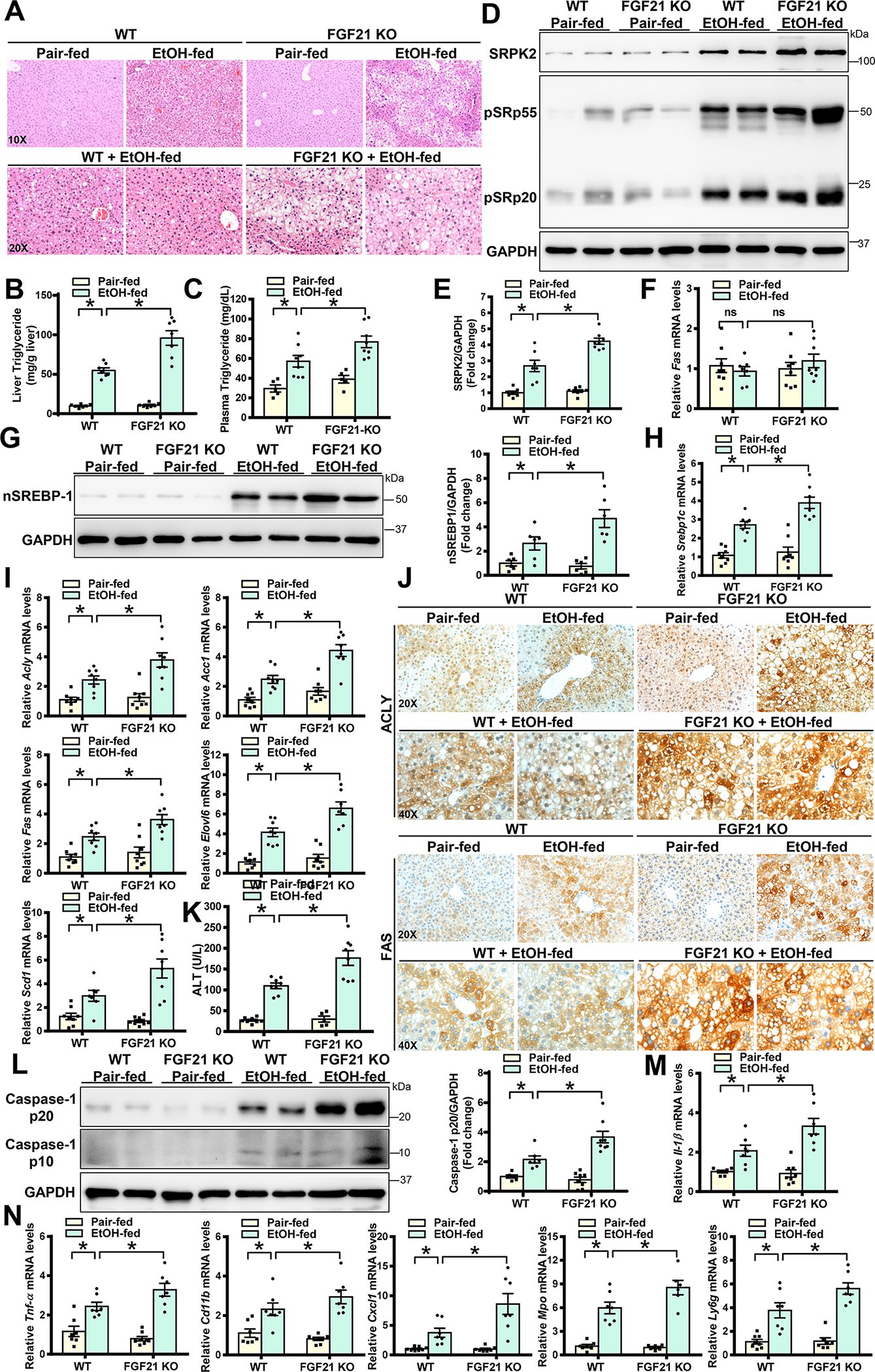
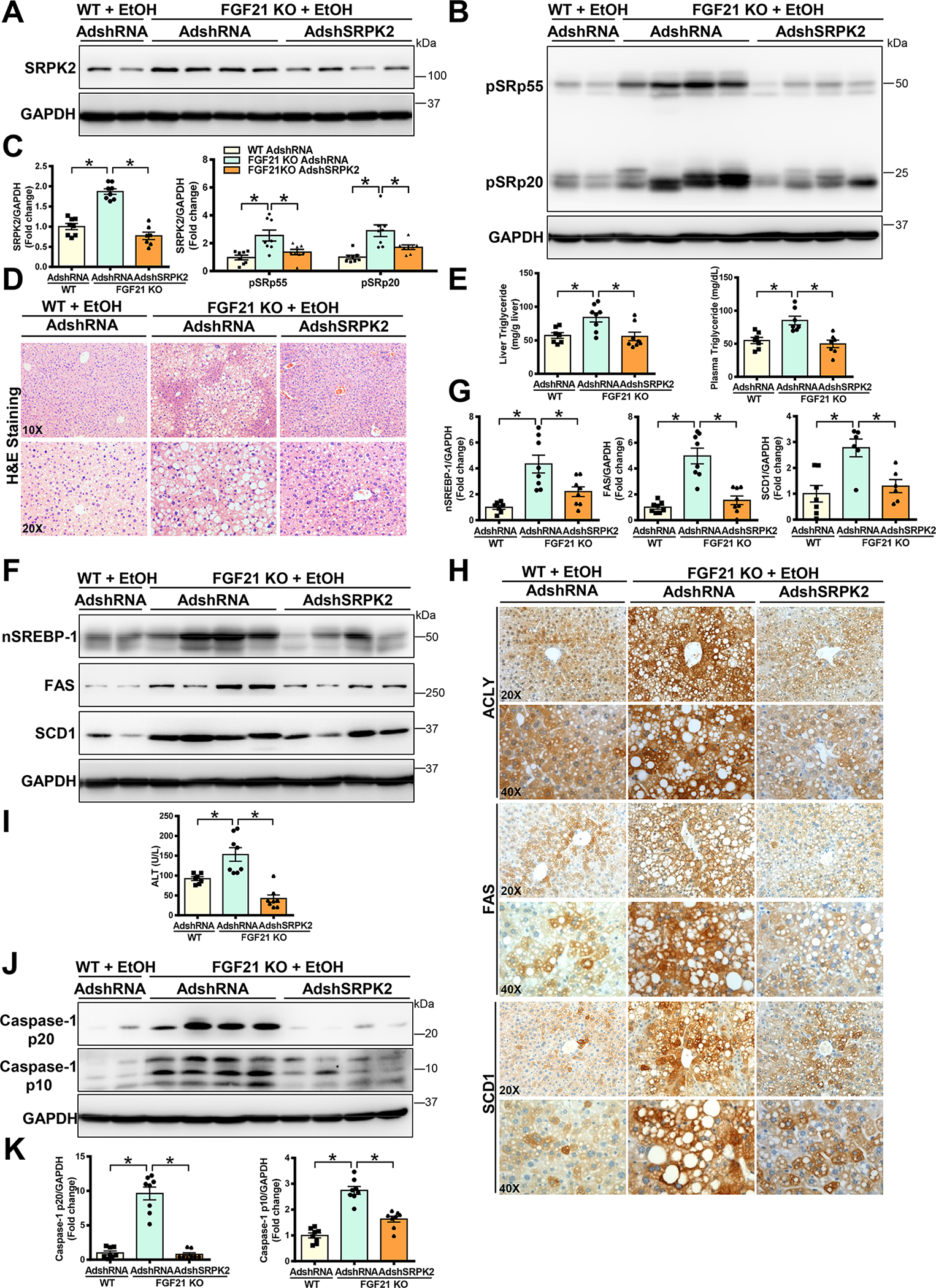
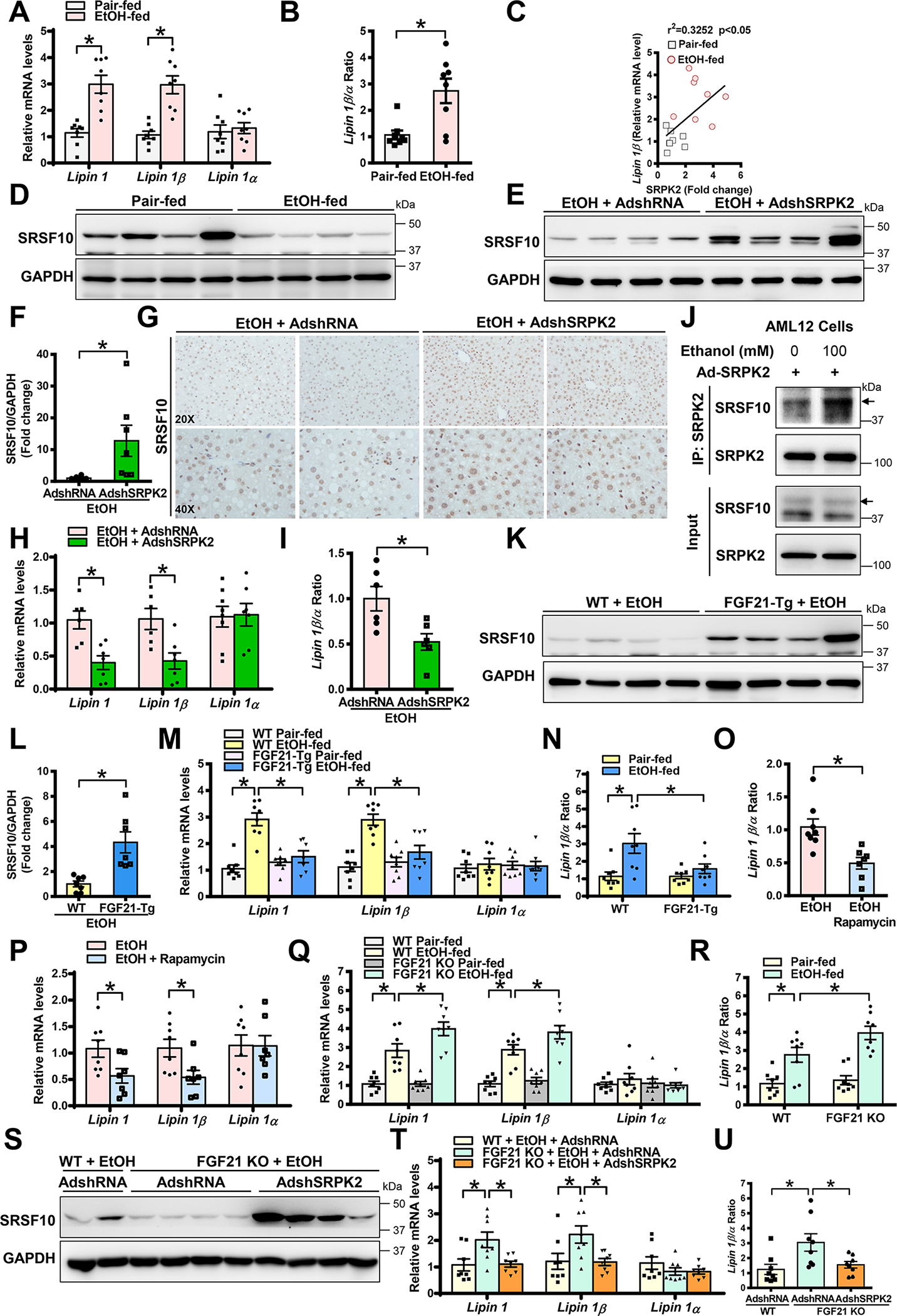
Approach and results: Serine-arginine-rich protein kinase 2 (SRPK2), a key kinase controlling alternative splicing, is activated in hepatocytes in response to alcohol, in mice with chronic-plus-binge alcohol feeding, and in patients with ALD. Such induction activates sterol regulatory element-binding protein 1 and promotes lipogenesis in ALD. Overexpression of FGF21 in transgenic mice abolishes alcohol-mediated induction of SRPK2 and its associated steatosis, lipotoxicity, and inflammation; these alcohol-induced pathologies are exacerbated in FGF21 knockout mice. Mechanistically, SRPK2 is required for alcohol-mediated impairment of serine-arginine splicing factor 10, which generates exon 7 inclusion in lipin 1 and triggers concurrent induction of lipogenic regulators-lipin 1β and sterol regulatory element-binding protein 1. FGF21 suppresses alcohol-induced SRPK2 accumulation through mammalian target of rapamycin complex 1 inhibition-dependent degradation of SRPK2. Silencing SRPK2 rescues alcohol-induced splicing dysregulation and liver injury in FGF21 knockout mice.
Conclusions: These studies reveal that (1) the regulation of alternative splicing by SRPK2 is implicated in lipogenesis in humans with ALD; (2) FGF21 is a key hepatokine that ameliorates ALD pathologies largely by inhibiting SRPK2; and (3) targeting SRPK2 signaling by FGF21 may offer potential therapeutic approaches to combat ALD.
Copyright © 2023 American Association for the Study of Liver Diseases.
Figures
Comment in
-
The pre-mRNA alternative splicing regulated by SRPK2: A new player in alcohol-associated liver disease?Hepatology. 2023 Nov 1;78(5):1329-1331. doi: 10.1097/HEP.0000000000000455. Epub 2023 May 16. Hepatology. 2023. PMID: 37183907 Free PMC article. No abstract available.
References
-
- Seitz HK, Bataller R, Cortez-Pinto H, Gao B, Gual A, Lackner C, Mathurin P, et al. Alcoholic liver disease. Nat Rev Dis Primers 2018;4:16. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
