

An Sfi1-like centrin-interacting centriolar plaque protein affects nuclear microtubule homeostasis
- PMID: 37130129
- PMCID: PMC10180636
- DOI: 10.1371/journal.ppat.1011325
An Sfi1-like centrin-interacting centriolar plaque protein affects nuclear microtubule homeostasis
Abstract
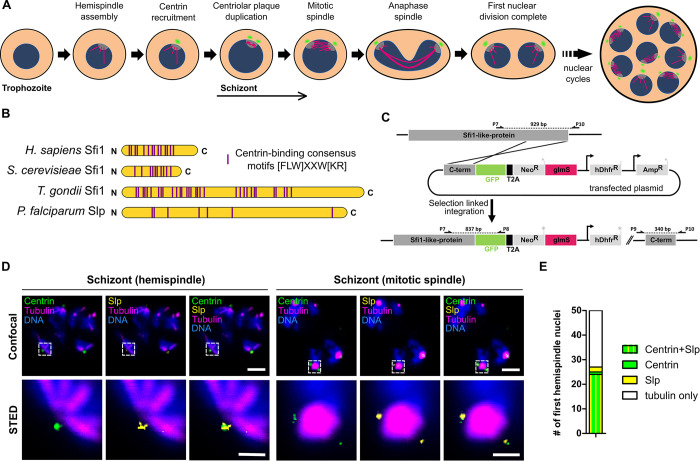
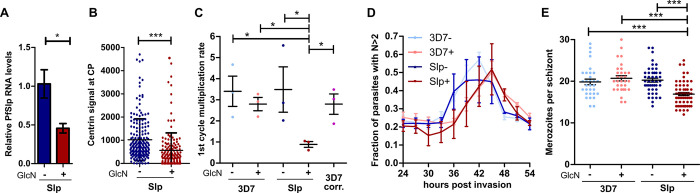
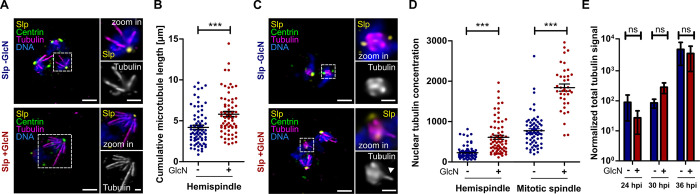
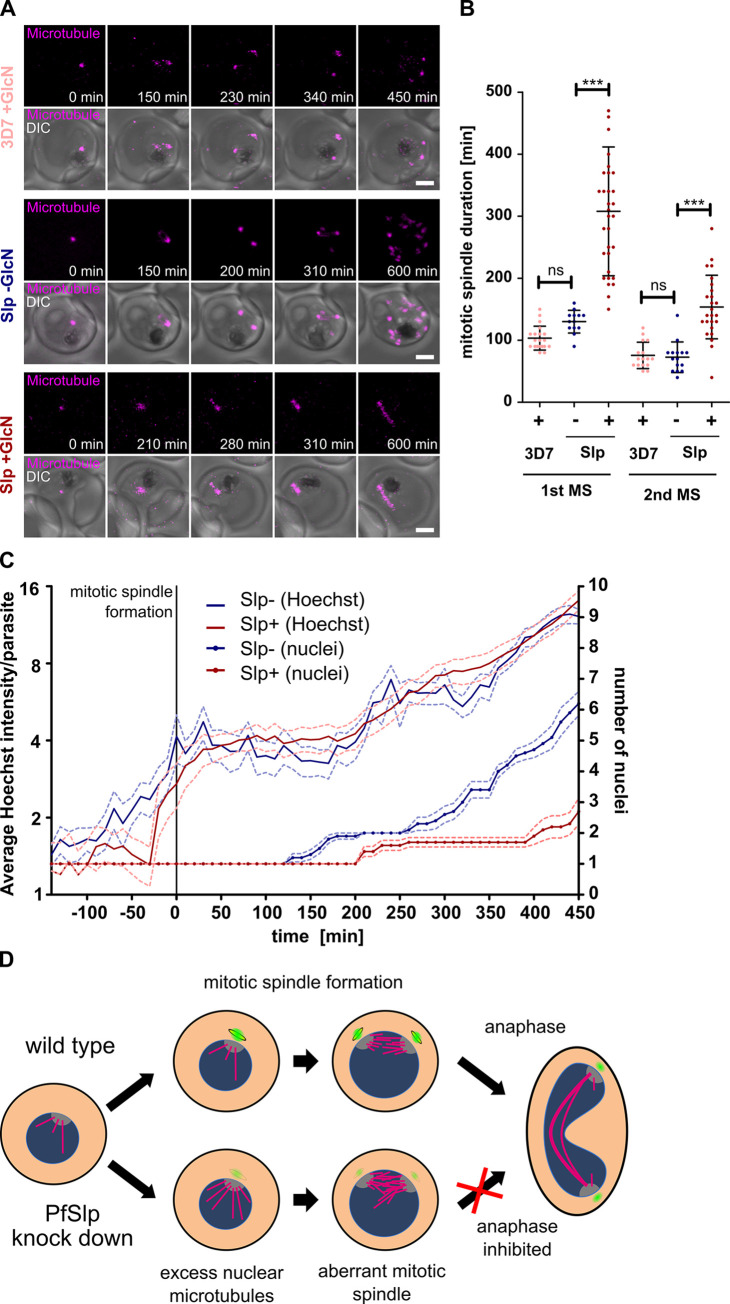
Malaria-causing parasites achieve rapid proliferation in human blood through multiple rounds of asynchronous nuclear division followed by daughter cell formation. Nuclear divisions critically depend on the centriolar plaque, which organizes intranuclear spindle microtubules. The centriolar plaque consists of an extranuclear compartment, which is connected via a nuclear pore-like structure to a chromatin-free intranuclear compartment. Composition and function of this non-canonical centrosome remain largely elusive. Centrins, which reside in the extranuclear part, are among the very few centrosomal proteins conserved in Plasmodium falciparum. Here we identify a novel centrin-interacting centriolar plaque protein. Conditional knock down of this Sfi1-like protein (PfSlp) caused a growth delay in blood stages, which correlated with a reduced number of daughter cells. Surprisingly, intranuclear tubulin abundance was significantly increased, which raises the hypothesis that the centriolar plaque might be implicated in regulating tubulin levels. Disruption of tubulin homeostasis caused excess microtubules and aberrant mitotic spindles. Time-lapse microscopy revealed that this prevented or delayed mitotic spindle extension but did not significantly interfere with DNA replication. Our study thereby identifies a novel extranuclear centriolar plaque factor and establishes a functional link to the intranuclear compartment of this divergent eukaryotic centrosome.
Copyright: © 2023 Wenz et al. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
References
-
- WHO. WHO Global, World Malaria Report 2021. Word Malaria report Geneva: World Health Organization. (2021). Licence: CC. 2021.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
