

This is a preprint.
A DRG genetic toolkit reveals molecular, morphological, and functional diversity of somatosensory neuron subtypes
- PMID: 37131664
- PMCID: PMC10153270
- DOI: 10.1101/2023.04.22.537932
A DRG genetic toolkit reveals molecular, morphological, and functional diversity of somatosensory neuron subtypes
Update in
-
A mouse DRG genetic toolkit reveals morphological and physiological diversity of somatosensory neuron subtypes.Cell. 2024 Mar 14;187(6):1508-1526.e16. doi: 10.1016/j.cell.2024.02.006. Epub 2024 Mar 4. Cell. 2024. PMID: 38442711 Free PMC article.
Abstract
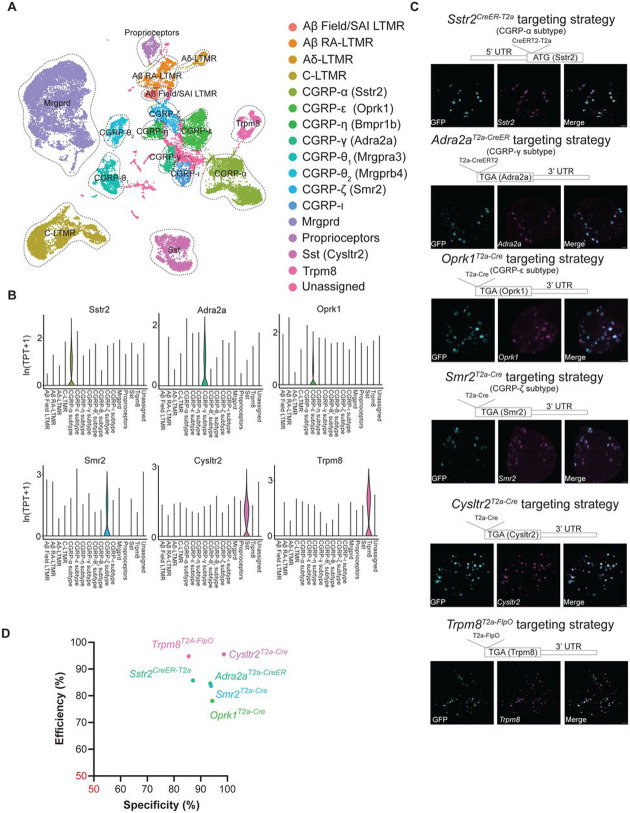
Mechanical and thermal stimuli acting on the skin are detected by morphologically and physiologically distinct sensory neurons of the dorsal root ganglia (DRG). Achieving a holistic view of how this diverse neuronal population relays sensory information from the skin to the central nervous system (CNS) has been challenging with existing tools. Here, we used transcriptomic datasets of the mouse DRG to guide development and curation of a genetic toolkit to interrogate transcriptionally defined DRG neuron subtypes. Morphological analysis revealed unique cutaneous axon arborization areas and branching patterns of each subtype. Physiological analysis showed that subtypes exhibit distinct thresholds and ranges of responses to mechanical and/or thermal stimuli. The somatosensory neuron toolbox thus enables comprehensive phenotyping of most principal sensory neuron subtypes. Moreover, our findings support a population coding scheme in which the activation thresholds of morphologically and physiologically distinct cutaneous DRG neuron subtypes tile multiple dimensions of stimulus space.
Figures






References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources