

The Efficiency of p27 Cleavage during In Vitro Respiratory Syncytial Virus (RSV) Infection Is Cell Line and RSV Subtype Dependent
- PMID: 37133390
- PMCID: PMC10231215
- DOI: 10.1128/jvi.00254-23
The Efficiency of p27 Cleavage during In Vitro Respiratory Syncytial Virus (RSV) Infection Is Cell Line and RSV Subtype Dependent
Abstract
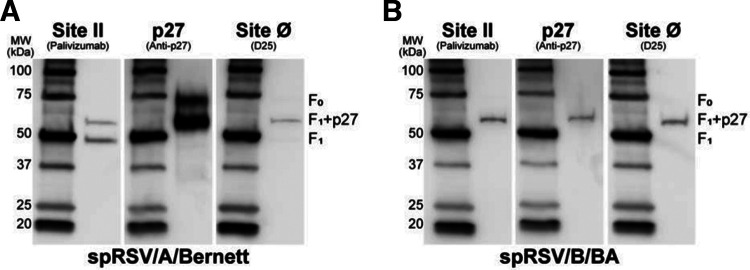
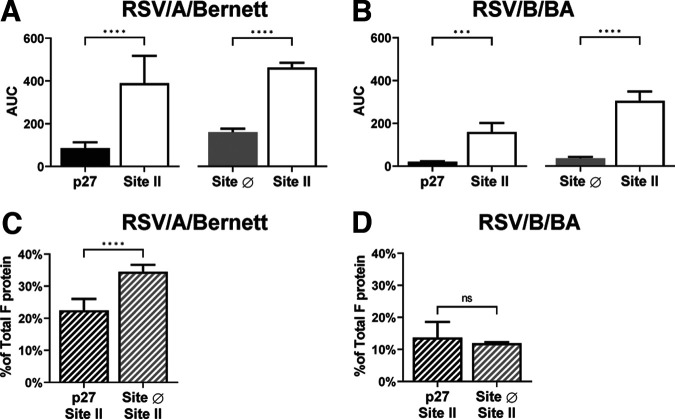
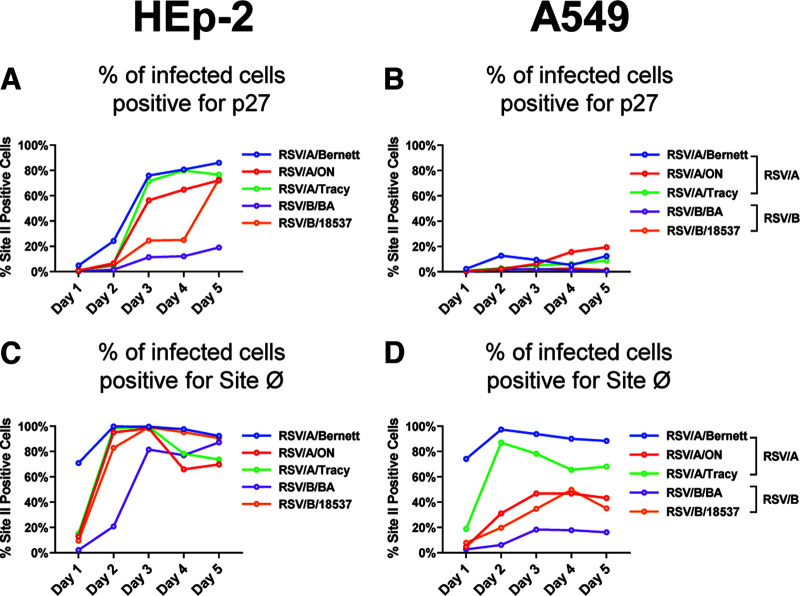
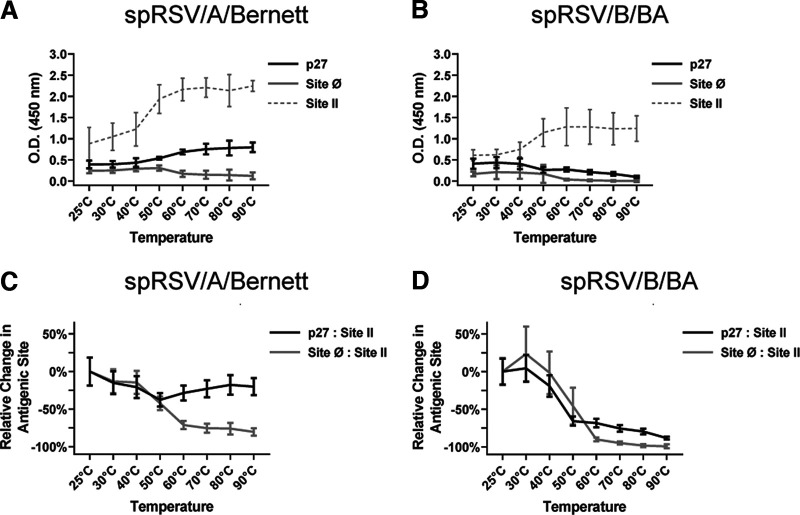
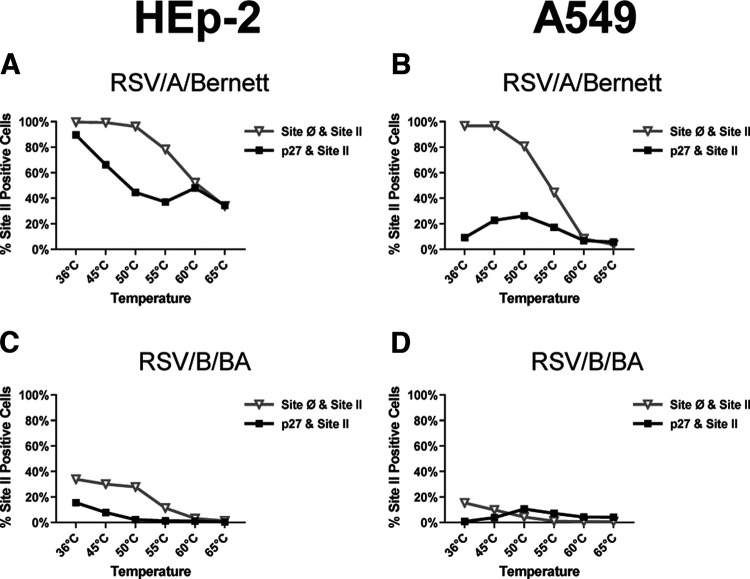
Respiratory syncytial virus (RSV) fusion protein (F) is highly conserved between subtypes A and B (RSV/A and RSV/B). To become fully active, F precursor undergoes enzymatic cleavage to yield F1 and F2 subunits and releases a 27-amino-acid peptide (p27). Virus-cell fusion occurs when RSV F undergoes a conformational change from pre-F to post-F. Previous data show that p27 is detected on RSV F, but questions remain regarding if and how p27 affects the conformation of mature RSV F. Monoclonal antibodies against p27, site Ø (pre-F specific), and site II were used to monitor RSV F conformation by enzyme-linked immunosorbent assay (ELISA) and imaging flow cytometry. Pre-F to post-F conformational change was induced by a temperature stress test. We found that p27 cleavage efficiency was lower on sucrose-purified RSV/A (spRSV/A) than on spRSV/B. In addition, cleavage of RSV F was cell line dependent: HEp-2 cells had higher retention of p27 than did A549 cells when infected with RSV. Higher levels of p27 were also found on RSV/A-infected cells than on RSV/B-infected cells. We observed that RSV/A F with higher p27 levels could better sustain the pre-F conformation during the temperature stress challenge in both spRSV- and RSV-infected cell lines. Our findings suggest that despite F sequence similarity, p27 of RSV subtypes was cleaved with different efficiencies, which were also dependent on the cell lines used for infection. Importantly, the presence of p27 was associated with greater stability of the pre-F conformation, supporting the possibility that RSV has more than one mechanism for fusion to the host cell. IMPORTANCE RSV fusion protein (F) plays an important role in entry and viral fusion to the host cell. The F undergoes proteolytic cleavages releasing a 27-amino-acid peptide (p27) to become fully functional. The role of p27 in viral entry and the function of the partially cleaved F containing p27 has been overlooked. p27 is thought to destabilize the F trimers, and thus, there is need for a fully cleaved F. In this study, we detected p27 on purified RSV virions and on the surface of virus-infected HEp-2 and A549 cells for circulating RSV strains of both subtypes. Higher levels of partially cleaved F containing p27 better sustained the pre-F conformation during the temperature stress challenge. Our findings highlight that the cleavage efficiency of p27 is different between RSV subtypes and among cell lines and that the presence of p27 contributes to the stability of the pre-F conformation.
Keywords: F protein; RSV; RSV subtypes; cleavage; p27; peptide 27; prefusion conformation; respiratory syncytial virus.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Antibody responses of healthy adults to the p27 peptide of respiratory syncytial virus fusion protein.Vaccine. 2022 Jan 24;40(3):536-543. doi: 10.1016/j.vaccine.2021.11.087. Epub 2021 Dec 10. Vaccine. 2022. PMID: 34903371 Free PMC article.
-
Respiratory syncytial virus prefusogenic fusion (F) protein nanoparticle vaccine: Structure, antigenic profile, immunogenicity, and protection.Vaccine. 2019 Sep 24;37(41):6112-6124. doi: 10.1016/j.vaccine.2019.07.089. Epub 2019 Aug 12. Vaccine. 2019. PMID: 31416644
-
Antigenicity and immunogenicity of unique prefusion-mimic F proteins presented on enveloped virus-like particles.Vaccine. 2019 Oct 16;37(44):6656-6664. doi: 10.1016/j.vaccine.2019.09.041. Epub 2019 Sep 18. Vaccine. 2019. PMID: 31542260 Free PMC article.
-
Clinical Potential of Prefusion RSV F-specific Antibodies.Trends Microbiol. 2018 Mar;26(3):209-219. doi: 10.1016/j.tim.2017.09.009. Epub 2017 Oct 17. Trends Microbiol. 2018. PMID: 29054341 Review.
-
[Prevention of respiratory syncytial virus infection in infants. What has been done and where are we today?].Andes Pediatr. 2023 Dec;94(6):672-680. doi: 10.32641/andespediatr.v94i6.4861. Andes Pediatr. 2023. PMID: 38329302 Review. Spanish.
Cited by
-
The RSV F p27 peptide: current knowledge, important questions.Front Microbiol. 2023 Jun 21;14:1219846. doi: 10.3389/fmicb.2023.1219846. eCollection 2023. Front Microbiol. 2023. PMID: 37415824 Free PMC article. Review.
-
Actin cytoskeleton remodeling disrupts physical barriers to infection and presents entry receptors to respiratory syncytial virus.J Gen Virol. 2023 Nov;104(11):001923. doi: 10.1099/jgv.0.001923. J Gen Virol. 2023. PMID: 38015055 Free PMC article.
-
Evaluating the Impact of N-Glycan Sequon Removal in the p27 Peptide on RSV F Protein Immunogenicity and Functionality.Viruses. 2024 Nov 28;16(12):1848. doi: 10.3390/v16121848. Viruses. 2024. PMID: 39772158 Free PMC article.
-
A single amino acid mutation alters multiple neutralization epitopes in the respiratory syncytial virus fusion glycoprotein.Npj Viruses. 2025 Apr 22;3(1):33. doi: 10.1038/s44298-025-00119-8. Npj Viruses. 2025. PMID: 40295799 Free PMC article.
-
Respiratory syncytial virus fuses with plasma membrane to infect primary cultures of bronchial epithelial cells.Front Microbiol. 2025 Feb 26;16:1498955. doi: 10.3389/fmicb.2025.1498955. eCollection 2025. Front Microbiol. 2025. PMID: 40099186 Free PMC article.
References
-
- Shi T, McAllister DA, O’Brien KL, Simoes EAF, Madhi SA, Gessner BD, Polack FP, Balsells E, Acacio S, Aguayo C, Alassani I, Ali A, Antonio M, Awasthi S, Awori JO, Azziz-Baumgartner E, Baggett HC, Baillie VL, Balmaseda A, Barahona A, Basnet S, Bassat Q, Basualdo W, Bigogo G, Bont L, Breiman RF, Brooks WA, Broor S, Bruce N, Bruden D, Buchy P, Campbell S, Carosone-Link P, Chadha M, Chipeta J, Chou M, Clara W, Cohen C, de Cuellar E, Dang D-A, Dash-Yandag B, Deloria-Knoll M, Dherani M, Eap T, Ebruke BE, Echavarria M, de Freitas Lázaro Emediato CC, Fasce RA, Feikin DR, Feng L, et al.. 2017. Global, regional, and national disease burden estimates of acute lower respiratory infections due to respiratory syncytial virus in young children in 2015: a systematic review and modelling study. Lancet 390:946–958. doi:10.1016/S0140-6736(17)30938-8. - DOI - PMC - PubMed
-
- Mazur NI, Löwensteyn YN, Willemsen JE, Gill CJ, Forman L, Mwananyanda LM, Blau DM, Breiman RF, Madhi SA, Mahtab S, Gurley ES, El Arifeen S, Assefa N, Scott JAG, Onyango D, Tippet Barr BA, Kotloff KL, Sow SO, Mandomando I, Ogbuanu I, Jambai A, Bassat Q, CHAMPS Network the RSV GOLD Study Group, Caballero MT, Polack FP, Omer S, Momin Kazi A, Simões EAF, Satav A, Bont LJ. 2021. Global respiratory syncytial virus–related infant community deaths. Clin Infect Dis 73:S229–S237. doi:10.1093/cid/ciab528. - DOI - PMC - PubMed
-
- Li Y, Reeves RM, Wang X, Bassat Q, Brooks WA, Cohen C, Moore DP, Nunes M, Rath B, Campbell H, Nair H, RSV Global Epidemiology Network, RESCEU investigators . 2019. Global patterns in monthly activity of influenza virus, respiratory syncytial virus, parainfluenza virus, and metapneumovirus: a systematic analysis. Lancet Glob Health 7:e1031–e1045. doi:10.1016/S2214-109X(19)30264-5. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
