

Pathophysiology of Dyt1- Tor1a dystonia in mice is mediated by spinal neural circuit dysfunction
- PMID: 37134150
- PMCID: PMC7614689
- DOI: 10.1126/scitranslmed.adg3904
Pathophysiology of Dyt1- Tor1a dystonia in mice is mediated by spinal neural circuit dysfunction
Abstract
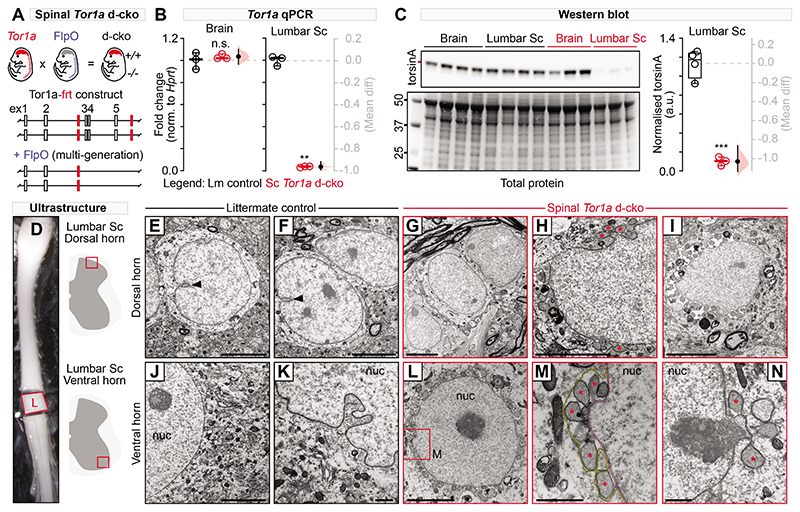
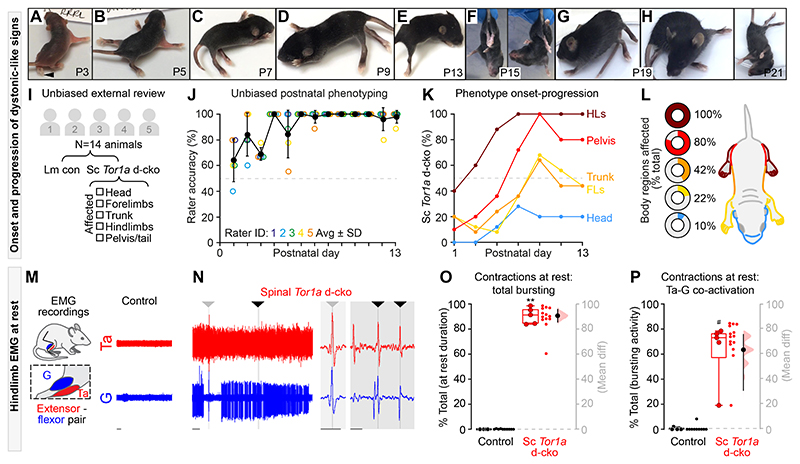
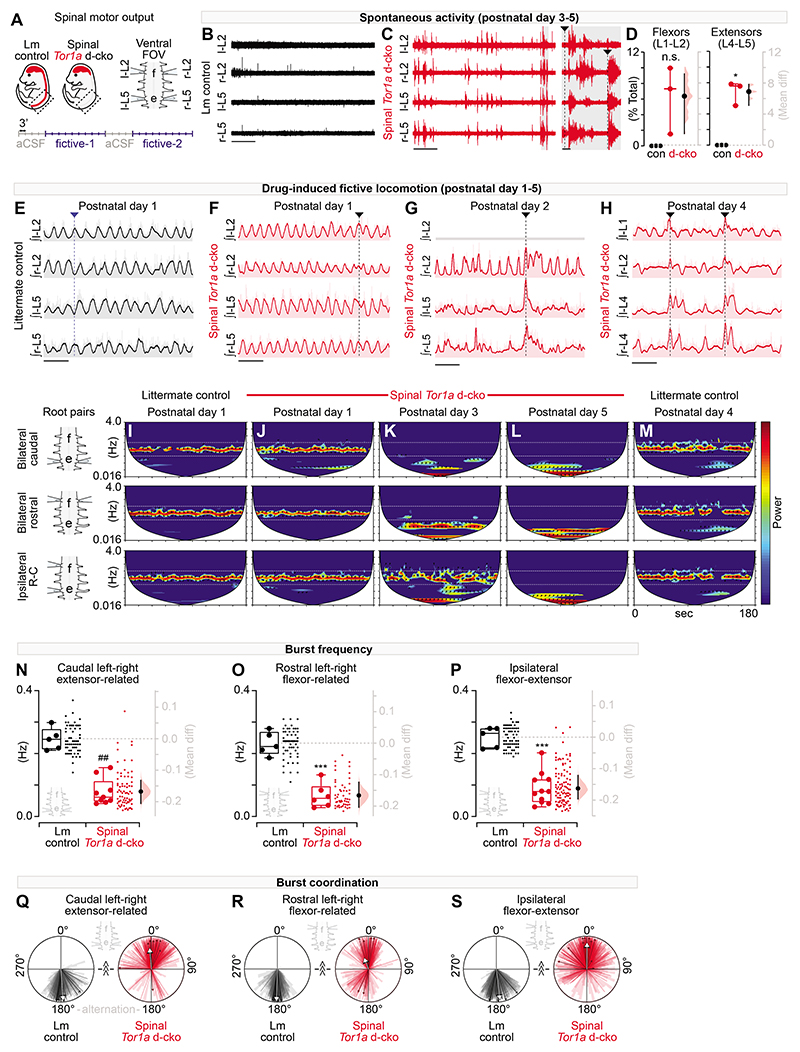
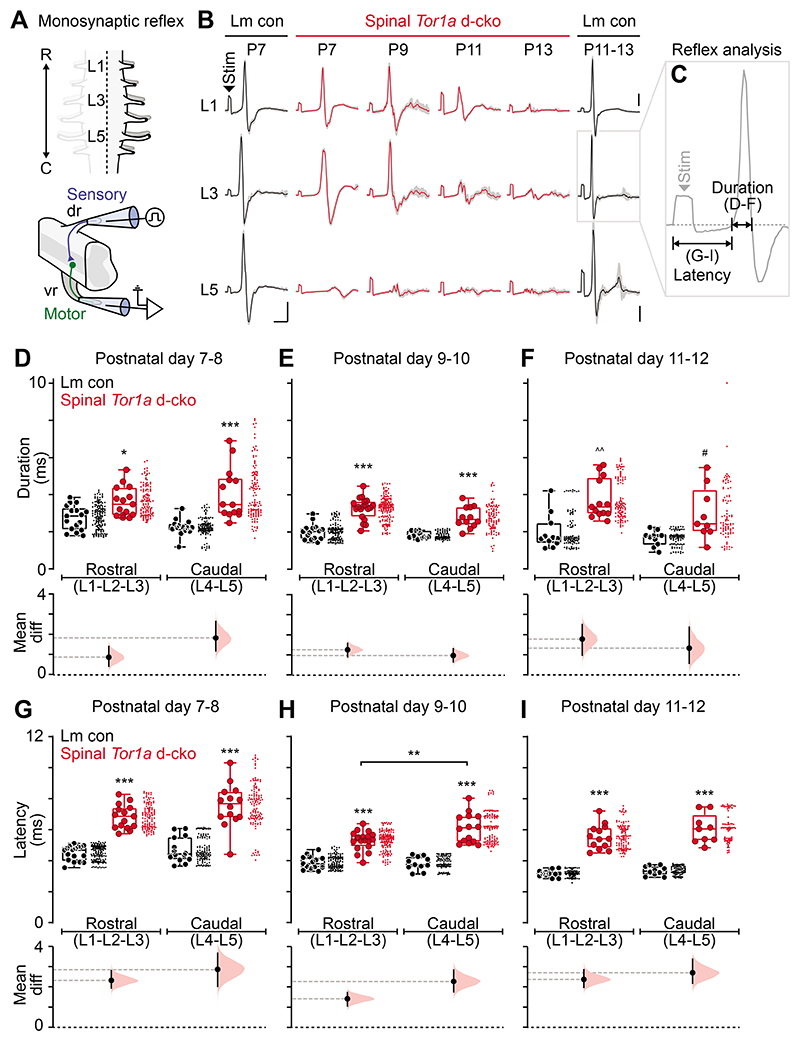
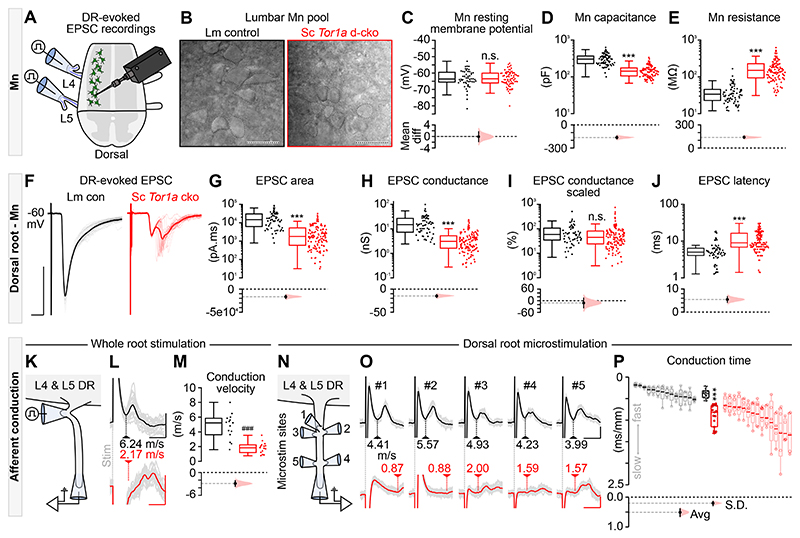
Dystonia, a neurological disorder defined by abnormal postures and disorganized movements, is considered to be a neural circuit disorder with dysfunction arising within and between multiple brain regions. Given that spinal neural circuits constitute the final pathway for motor control, we sought to determine their contribution to this movement disorder. Focusing on the most common inherited form of dystonia in humans, DYT1-TOR1A, we generated a conditional knockout of the torsin family 1 member A (Tor1a) gene in the mouse spinal cord and dorsal root ganglia (DRG). We found that these mice recapitulated the phenotype of the human condition, developing early-onset generalized torsional dystonia. Motor signs emerged early in the mouse hindlimbs before spreading caudo-rostrally to affect the pelvis, trunk, and forelimbs throughout postnatal maturation. Physiologically, these mice bore the hallmark features of dystonia, including spontaneous contractions at rest and excessive and disorganized contractions, including cocontractions of antagonist muscle groups, during voluntary movements. Spontaneous activity, disorganized motor output, and impaired monosynaptic reflexes, all signs of human dystonia, were recorded from isolated mouse spinal cords from these conditional knockout mice. All components of the monosynaptic reflex arc were affected, including motor neurons. Given that confining the Tor1a conditional knockout to DRG did not lead to early-onset dystonia, we conclude that the pathophysiological substrate of this mouse model of dystonia lies in spinal neural circuits. Together, these data provide new insights into our current understanding of dystonia pathophysiology.
Conflict of interest statement
RMB is a co-founder and is on the board of Sania Therapeutics, Inc and consults for Sania Rx Ltd. CCS is an employee of Sania Rx, Ltd.
Figures
Similar articles
-
Decreased number of striatal cholinergic interneurons and motor deficits in dopamine receptor 2-expressing-cell-specific Dyt1 conditional knockout mice.Neurobiol Dis. 2020 Feb;134:104638. doi: 10.1016/j.nbd.2019.104638. Epub 2019 Oct 13. Neurobiol Dis. 2020. PMID: 31618684 Free PMC article.
-
Motor deficit and lack of overt dystonia in Dlx conditional Dyt1 knockout mice.Behav Brain Res. 2023 Feb 15;439:114221. doi: 10.1016/j.bbr.2022.114221. Epub 2022 Nov 20. Behav Brain Res. 2023. PMID: 36417958 Free PMC article.
-
Cell-specific effects of Dyt1 knock-out on sensory processing, network-level connectivity, and motor deficits.Exp Neurol. 2021 Sep;343:113783. doi: 10.1016/j.expneurol.2021.113783. Epub 2021 Jun 10. Exp Neurol. 2021. PMID: 34119482 Free PMC article.
-
Membrane defects and genetic redundancy: Are we at a turning point for DYT1 dystonia?Mov Disord. 2017 Mar;32(3):371-381. doi: 10.1002/mds.26880. Epub 2016 Dec 2. Mov Disord. 2017. PMID: 27911022 Review.
-
[Torsin 1A and the pathomechanism of torsion dystonia type 1].Postepy Biochem. 2015;61(1):35-41. Postepy Biochem. 2015. PMID: 26281352 Review. Polish.
Cited by
-
Definition and Classification of Dystonia.Mov Disord. 2025 Jul;40(7):1248-1259. doi: 10.1002/mds.30220. Epub 2025 May 6. Mov Disord. 2025. PMID: 40326714 Free PMC article. Review.
-
Spinal Dystonia Associated with Transverse Myelitis in an Adolescent Female: A Case Report.Adolesc Health Med Ther. 2024 Jan 20;15:1-4. doi: 10.2147/AHMT.S445606. eCollection 2024. Adolesc Health Med Ther. 2024. PMID: 38274981 Free PMC article.
-
Neurobiology of Dystonia: Review of Genetics, Animal Models, and Neuroimaging.Brain Sci. 2025 Jul 19;15(7):767. doi: 10.3390/brainsci15070767. Brain Sci. 2025. PMID: 40722357 Free PMC article. Review.
-
Air-stepping in the neonatal mouse: a powerful tool for analyzing early stages of rhythmic limb movement development.J Neurophysiol. 2024 Feb 1;131(2):321-337. doi: 10.1152/jn.00227.2023. Epub 2024 Jan 10. J Neurophysiol. 2024. PMID: 38198656 Free PMC article.
-
Aberrant outputs of glutamatergic neurons in deep cerebellar nuclei mediate dystonic movements.Sci Adv. 2025 May 9;11(19):eadp2377. doi: 10.1126/sciadv.adp2377. Epub 2025 May 9. Sci Adv. 2025. PMID: 40344058 Free PMC article.
References
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
