

Massively parallel base editing to map variant effects in human hematopoiesis
- PMID: 37137305
- PMCID: PMC10225359
- DOI: 10.1016/j.cell.2023.03.035
Massively parallel base editing to map variant effects in human hematopoiesis
Abstract
Systematic evaluation of the impact of genetic variants is critical for the study and treatment of human physiology and disease. While specific mutations can be introduced by genome engineering, we still lack scalable approaches that are applicable to the important setting of primary cells, such as blood and immune cells. Here, we describe the development of massively parallel base-editing screens in human hematopoietic stem and progenitor cells. Such approaches enable functional screens for variant effects across any hematopoietic differentiation state. Moreover, they allow for rich phenotyping through single-cell RNA sequencing readouts and separately for characterization of editing outcomes through pooled single-cell genotyping. We efficiently design improved leukemia immunotherapy approaches, comprehensively identify non-coding variants modulating fetal hemoglobin expression, define mechanisms regulating hematopoietic differentiation, and probe the pathogenicity of uncharacterized disease-associated variants. These strategies will advance effective and high-throughput variant-to-function mapping in human hematopoiesis to identify the causes of diverse diseases.
Keywords: base editing; differentiation; functional screens; genome engineering; hematopoiesis; hematopoietic stem cell; primary cells; single-cell genomics.
Copyright © 2023 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests D.R.L. and G.A.N. have filed patent applications on gene-editing technologies through the Broad Institute of MIT and Harvard. D.R.L. is a consultant and equity owner of Beam Therapeutics, Pairwise Plants, Prime Medicine, Chroma Medicine and Nvelop Therapeutics, companies that use or deliver genome editing or genome engineering technologies. R.J.X. is the co-founder of Jnana Therapeutics and Celsius Therapeutics, the director of Moonlake Immunotherapeutics and a scientific advisory board member to Nestle, all unrelated to the present work. V.G.S. serves as an advisor to and/or has equity in Branch Biosciences, Ensoma, Novartis, Forma, and Cellarity, all unrelated to the present work.
Figures






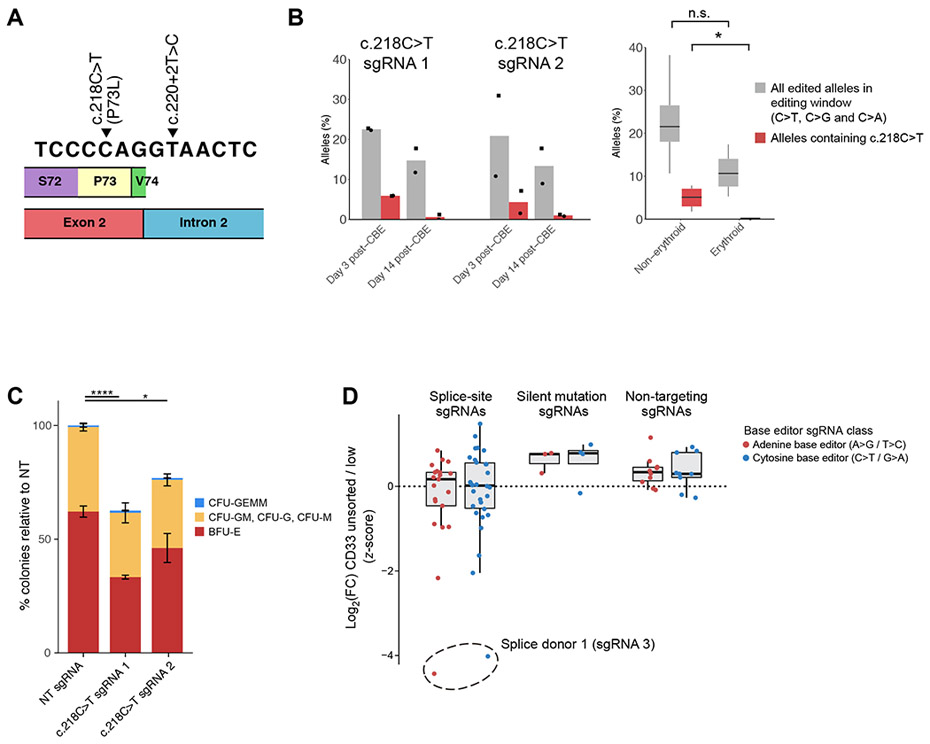
Comment in
-
Base-editing screens illuminate variant effects in human hematopoiesis.Cell Rep Methods. 2023 Jul 24;3(7):100541. doi: 10.1016/j.crmeth.2023.100541. eCollection 2023 Jul 24. Cell Rep Methods. 2023. PMID: 37533644 Free PMC article.
References
-
- Schiroli G, Conti A, Ferrari S, Della Volpe L, Jacob A, Albano L, Beretta S, Calabria A, Vavassori V, Gasparini P, et al. (2019). Precise Gene Editing Preserves Hematopoietic Stem Cell Function following Transient p53-Mediated DNA Damage Response. Cell Stem Cell 24, 551–565.e8. 10.1016/j.stem.2019.02.019. - DOI - PMC - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
