

Vaginal epithelial dysfunction is mediated by the microbiome, metabolome, and mTOR signaling
- PMID: 37149863
- PMCID: PMC10242450
- DOI: 10.1016/j.celrep.2023.112474
Vaginal epithelial dysfunction is mediated by the microbiome, metabolome, and mTOR signaling
Abstract
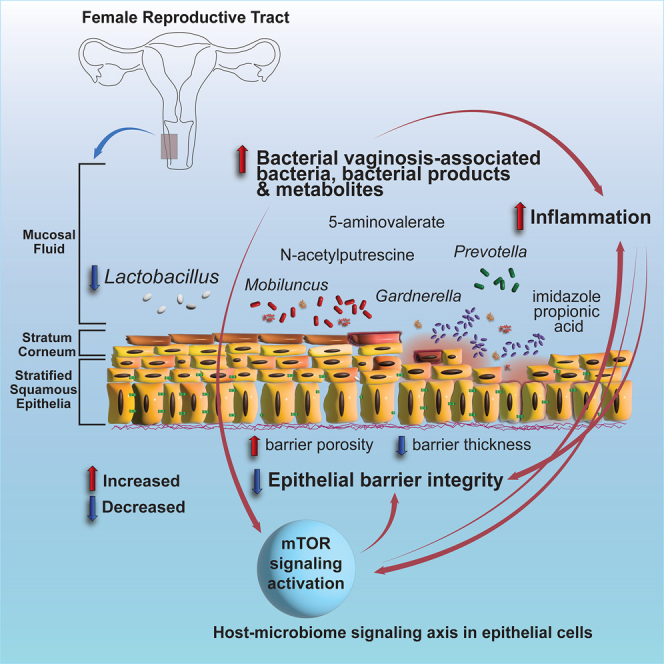
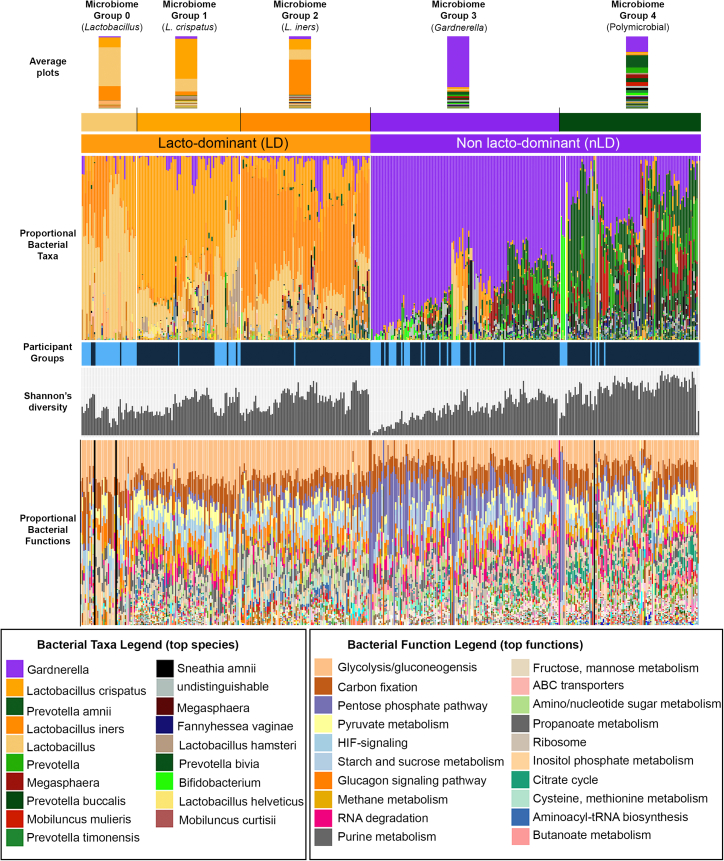
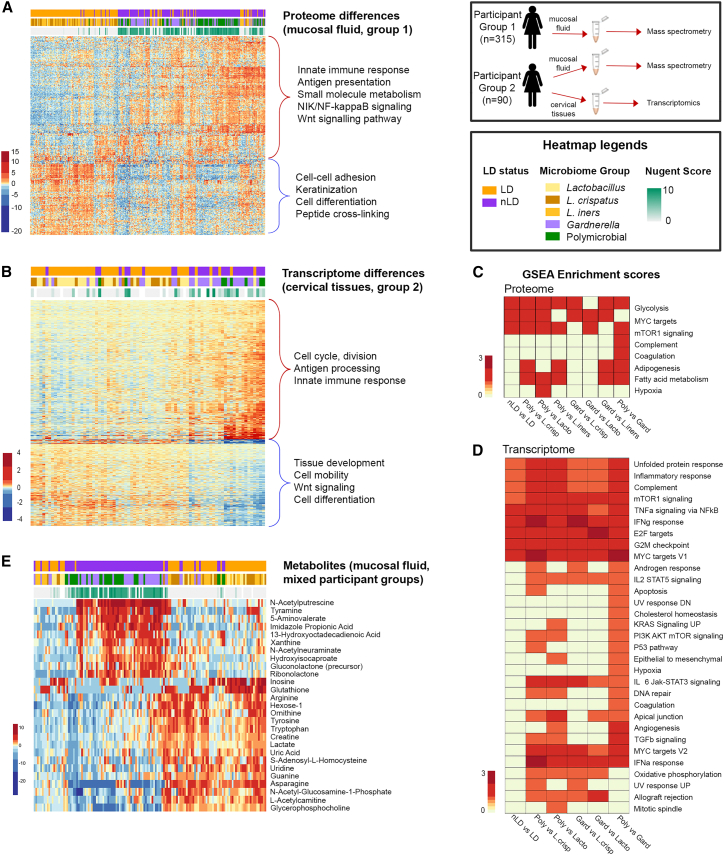
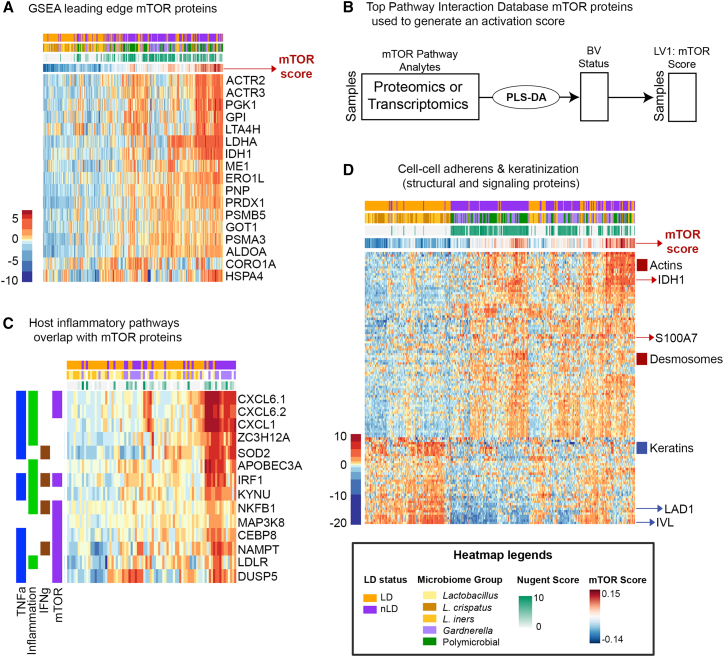
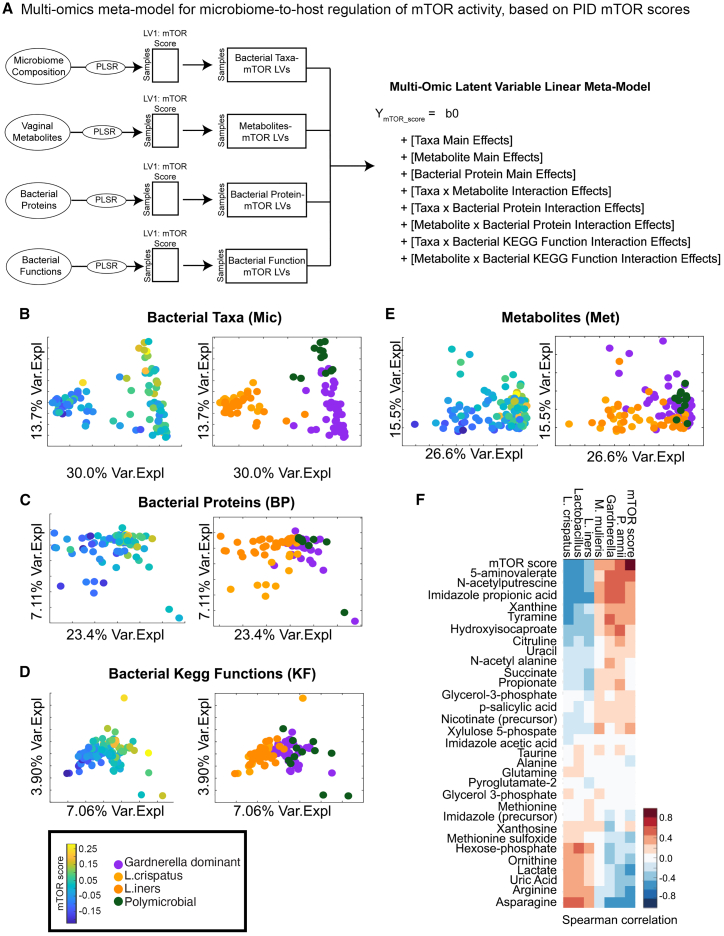
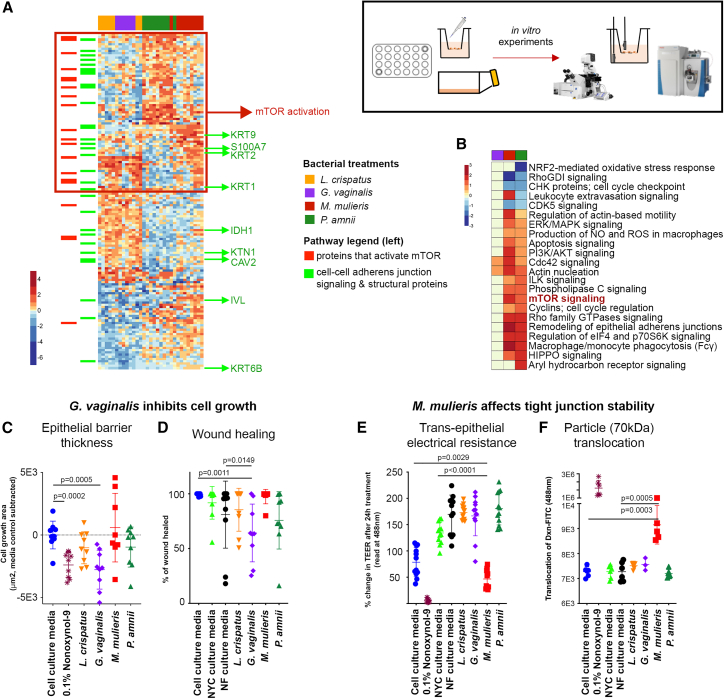
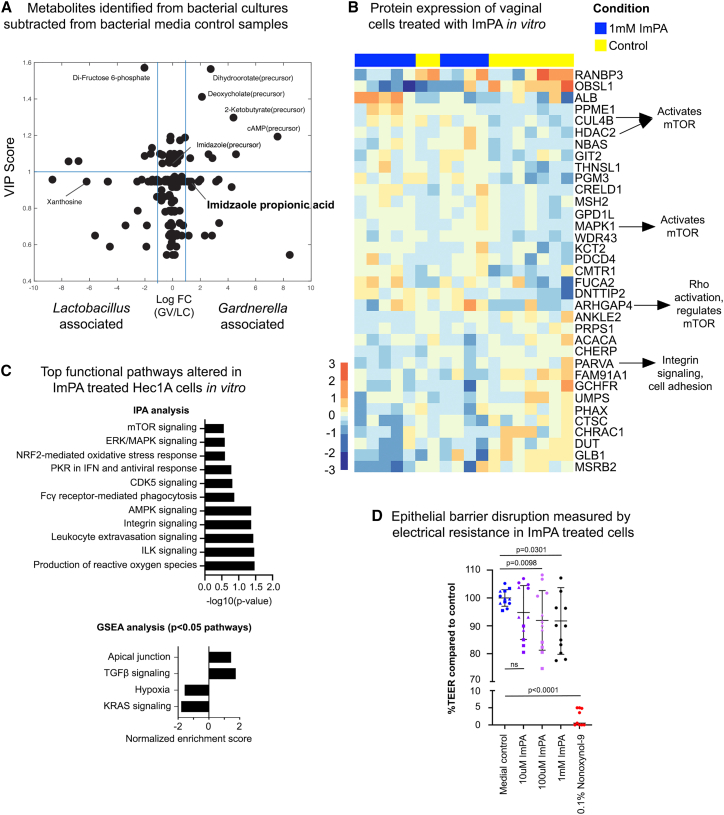
Bacterial vaginosis (BV) is characterized by depletion of Lactobacillus and overgrowth of anaerobic and facultative bacteria, leading to increased mucosal inflammation, epithelial disruption, and poor reproductive health outcomes. However, the molecular mediators contributing to vaginal epithelial dysfunction are poorly understood. Here we utilize proteomic, transcriptomic, and metabolomic analyses to characterize biological features underlying BV in 405 African women and explore functional mechanisms in vitro. We identify five major vaginal microbiome groups: L. crispatus (21%), L. iners (18%), Lactobacillus (9%), Gardnerella (30%), and polymicrobial (22%). Using multi-omics we show that BV-associated epithelial disruption and mucosal inflammation link to the mammalian target of rapamycin (mTOR) pathway and associate with Gardnerella, M. mulieris, and specific metabolites including imidazole propionate. Experiments in vitro confirm that type strain G. vaginalis and M. mulieris supernatants and imidazole propionate directly affect epithelial barrier function and activation of mTOR pathways. These results find that the microbiome-mTOR axis is a central feature of epithelial dysfunction in BV.
Keywords: BV; CP: Microbiology; bacterial vaginosis; epithelial barrier; imidazolepropionic acid; in vitro models; inflammation; mTOR signaling; metabolomics; proteomics; transcriptomics; vaginal dysbiosis; vaginal microbiome.
Copyright © 2023 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests J.M.B. is an employee of Gilead Sciences, outside of the submitted work.
Figures
References
-
- Anahtar M.N., Byrne E.H., Doherty K.E., Bowman B.A., Yamamoto H.S., Soumillon M., Padavattan N., Ismail N., Moodley A., Sabatini M.E., et al. Cervicovaginal bacteria are a major modulator of host inflammatory responses in the female genital tract. Immunity. 2015;42:965–976. doi: 10.1016/j.immuni.2015.04.019. - DOI - PMC - PubMed
-
- Aldunate M., Srbinovski D., Hearps A.C., Latham C.F., Ramsland P.A., Gugasyan R., Cone R.A., Tachedjian G. Antimicrobial and immune modulatory effects of lactic acid and short chain fatty acids produced by vaginal microbiota associated with eubiosis and bacterial vaginosis. Front. Physiol. 2015;6:164. doi: 10.3389/fphys.2015.00164. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
