

Convergence of YAP/TAZ, TEAD and TP63 activity is associated with bronchial premalignant severity and progression
- PMID: 37150829
- PMCID: PMC10165825
- DOI: 10.1186/s13046-023-02674-5
Convergence of YAP/TAZ, TEAD and TP63 activity is associated with bronchial premalignant severity and progression
Abstract
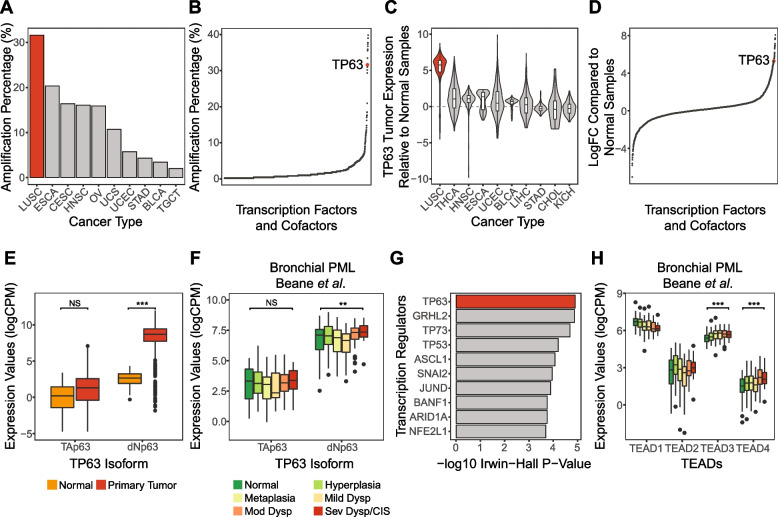
Background: Bronchial premalignant lesions (PMLs) are composed primarily of cells resembling basal epithelial cells of the airways, which through poorly understood mechanisms have the potential to progress to lung squamous cell carcinoma (LUSC). Despite ongoing efforts that have mapped gene expression and cell diversity across bronchial PML pathologies, signaling and transcriptional events driving malignancy are poorly understood. Evidence has suggested key roles for the Hippo pathway effectors YAP and TAZ and associated TEAD and TP63 transcription factor families in bronchial basal cell biology and LUSC. In this study we examine the functional association of YAP/TAZ, TEADs and TP63 in bronchial epithelial cells and PMLs.
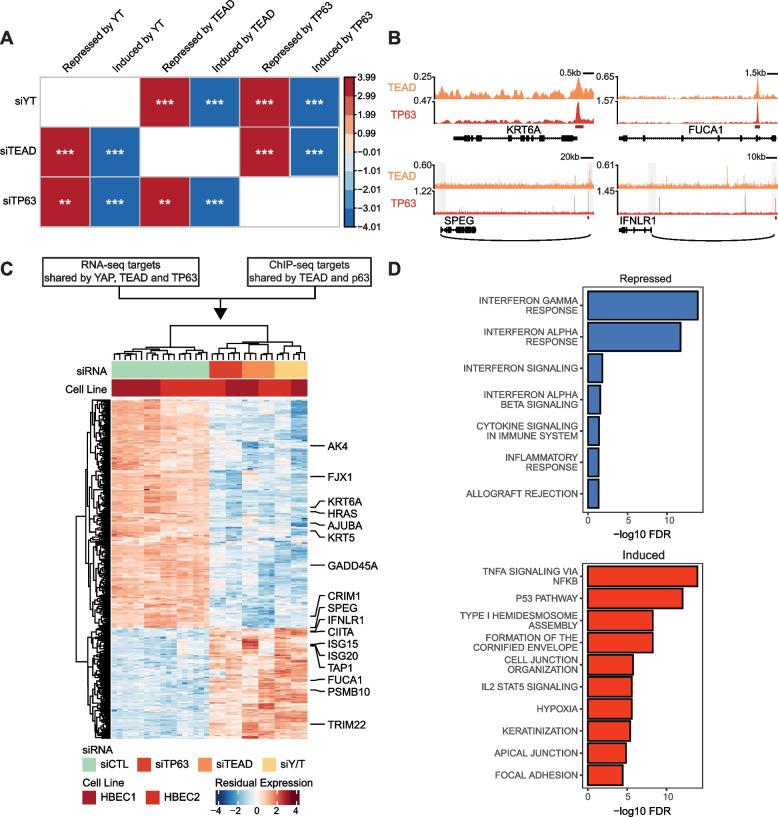
Methods: We performed RNA-seq in primary human bronchial epithelial cells following small interfering RNA (siRNA)-mediated depletion of YAP/TAZ, TEADs or TP63, and combined these data with ChIP-seq analysis of these factors. Directly activated or repressed genes were identified and overlapping genes were profiled across gene expression data obtained from progressive or regressive human PMLs and across lung single cell RNA-seq data sets.
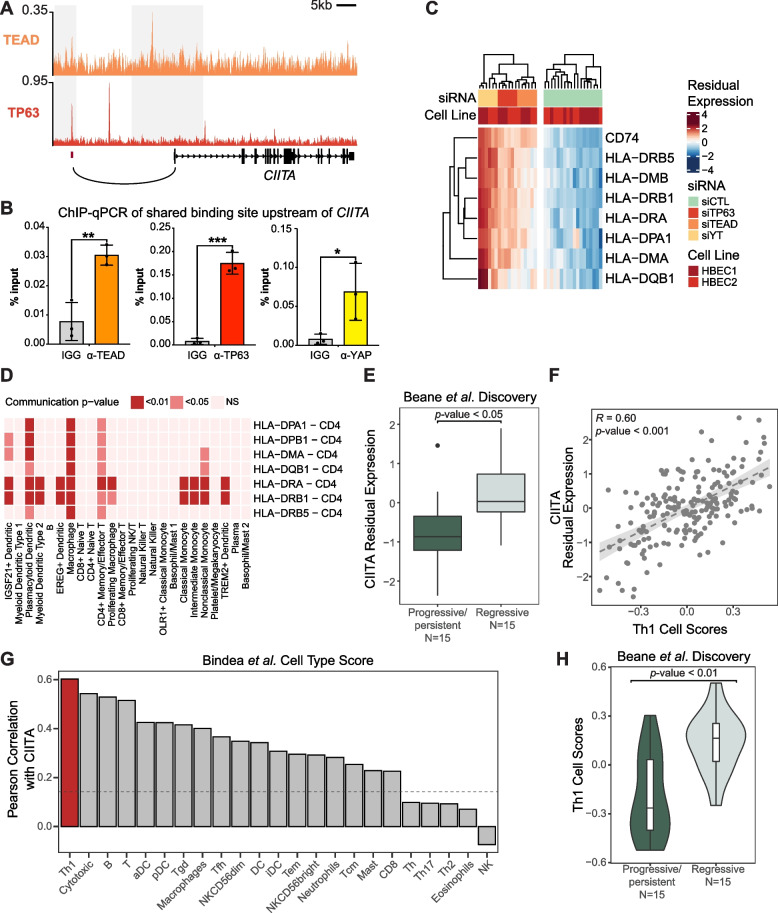
Results: Analysis of genes regulated by YAP/TAZ, TEADs, and TP63 in human bronchial epithelial cells revealed a converged transcriptional network that is strongly associated with the pathological progression of bronchial PMLs. Our observations suggest that YAP/TAZ-TEAD-TP63 associate to cooperatively promote basal epithelial cell proliferation and repress signals associated with interferon responses and immune cell communication. Directly repressed targets we identified include the MHC Class II transactivator CIITA, which is repressed in progressive PMLs and associates with adaptive immune responses in the lung. Our findings provide molecular insight into the control of gene expression events driving PML progression, including those contributing to immune evasion, offering potential new avenues for lung cancer interception.
Conclusions: Our study identifies important gene regulatory functions for YAP/TAZ-TEAD-TP63 in the early stages of lung cancer development, which notably includes immune-suppressive roles, and suggest that an assessment of the activity of this transcriptional complex may offer a means to identify immune evasive bronchial PMLs and serve as a potential therapeutic target.
Keywords: Bronchial premalignant lesions; CIITA; Immune evasion; MHCII; TAZ; TEAD; TP63; YAP.
© 2023. The Author(s).
Conflict of interest statement
X.V., S.M., M.E.L., and J.B. have received funding from sponsored research with Janssen Pharmaceuticals. Additionally, A.S. and M.E. are employees of Janssen Pharmaceuticals. All other authors declare no competing interests.
Figures



References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
