

This is a preprint.
Rit2 silencing in dopamine neurons drives a Parkinsonian phenotype
- PMID: 37162843
- PMCID: PMC10168302
- DOI: 10.1101/2023.04.26.538430
Rit2 silencing in dopamine neurons drives a Parkinsonian phenotype
Abstract
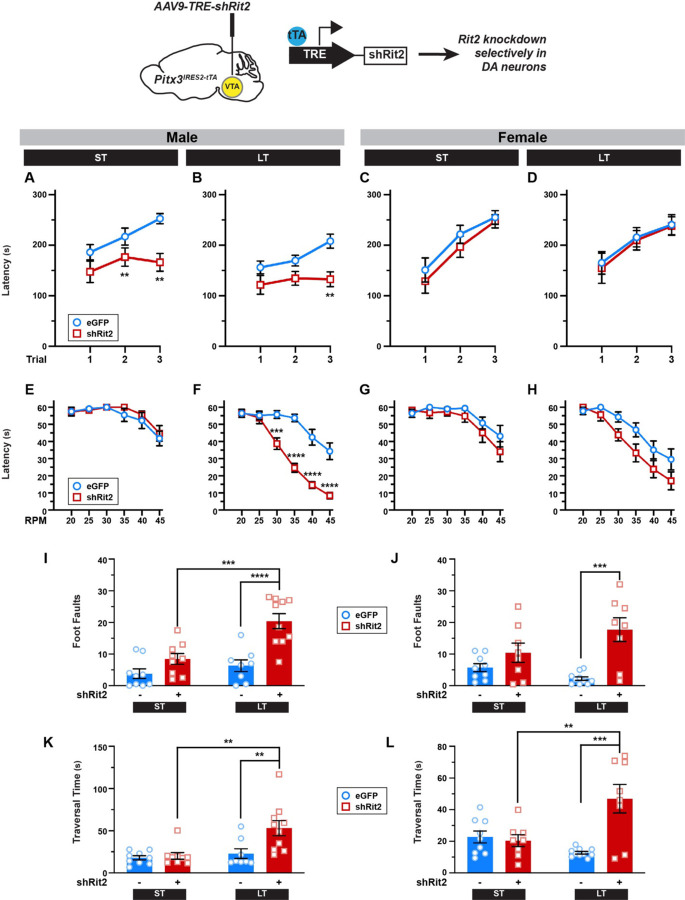
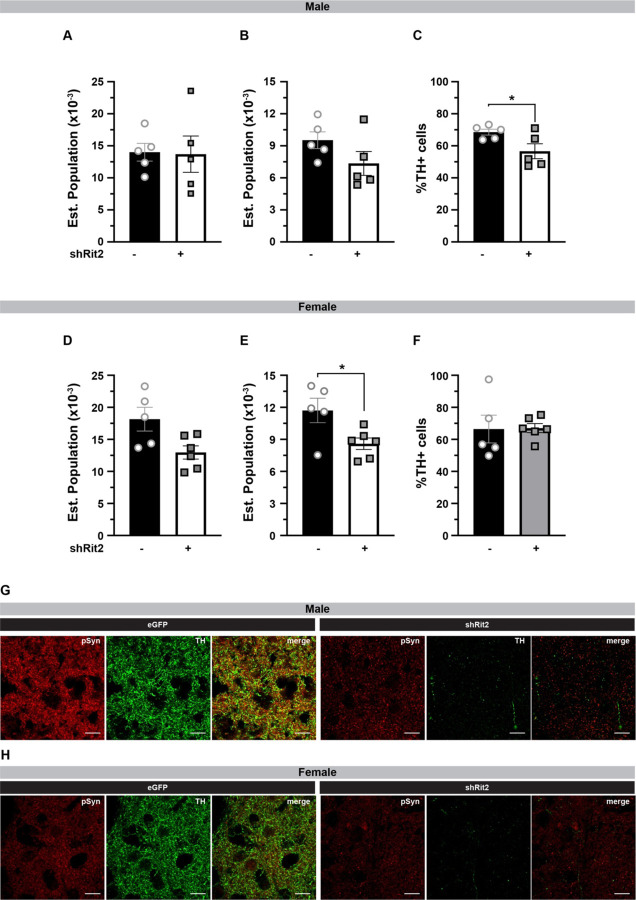
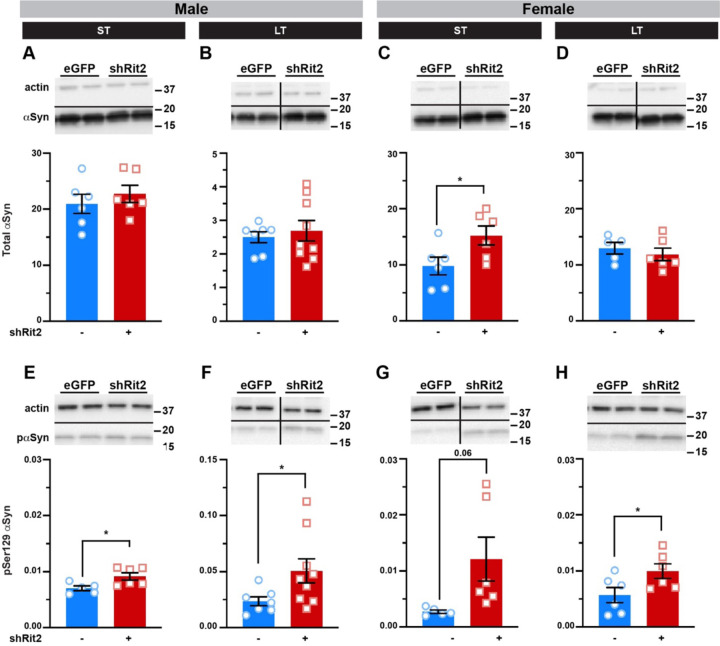
Parkinson's disease (PD) is the second most prevalent neurodegenerative disease and arises from dopamine (DA) neuron death selectively in the substantia nigra pars compacta (SNc). Rit2 is a reported PD risk allele, and recent single cell transcriptomic studies identified a major RIT2 cluster in PD DA neurons, potentially linking Rit2 expression loss to a PD patient cohort. However, it is still unknown whether Rit2 loss itself is causative for PD or PD-like symptoms. Here we report that conditional Rit2 silencing in mouse DA neurons drove motor dysfunction that occurred earlier in males than females and was rescued at early stages by either inhibiting the DA transporter (DAT) or with L-DOPA treatment. Motor dysfunction was accompanied by decreased DA release, striatal DA content, phenotypic DAergic markers, DA neurons, and DAergic terminals, with increased pSer129-alpha synuclein and pSer935-LRRK2 expression. These results provide the first evidence that Rit2 loss is causal for SNc cell death and a PD-like phenotype, and reveal key sex-specific differences in the response to Rit2 loss.
Conflict of interest statement
Conflict-of-interest statement The authors have declared that no conflict of interest exists.
Figures




References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous