

This is a preprint.
Assessment of three-dimensional RNA structure prediction in CASP15
- PMID: 37162955
- PMCID: PMC10168427
- DOI: 10.1101/2023.04.25.538330
Assessment of three-dimensional RNA structure prediction in CASP15
Update in
-
Assessment of three-dimensional RNA structure prediction in CASP15.Proteins. 2023 Dec;91(12):1747-1770. doi: 10.1002/prot.26602. Epub 2023 Oct 24. Proteins. 2023. PMID: 37876231 Free PMC article.
Abstract
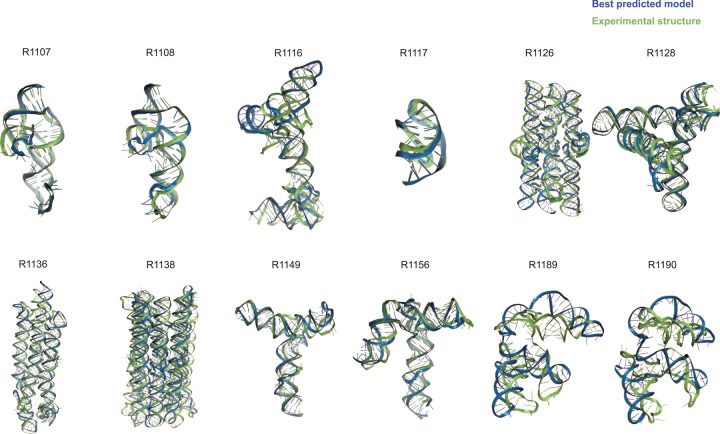
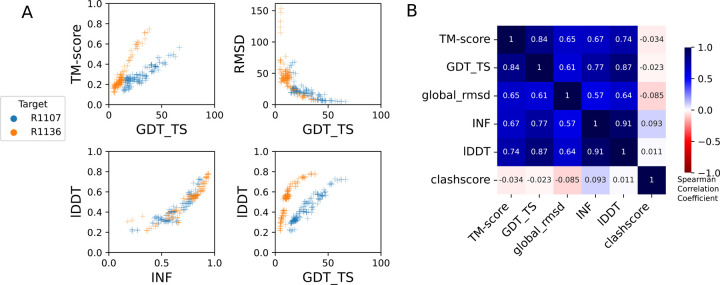
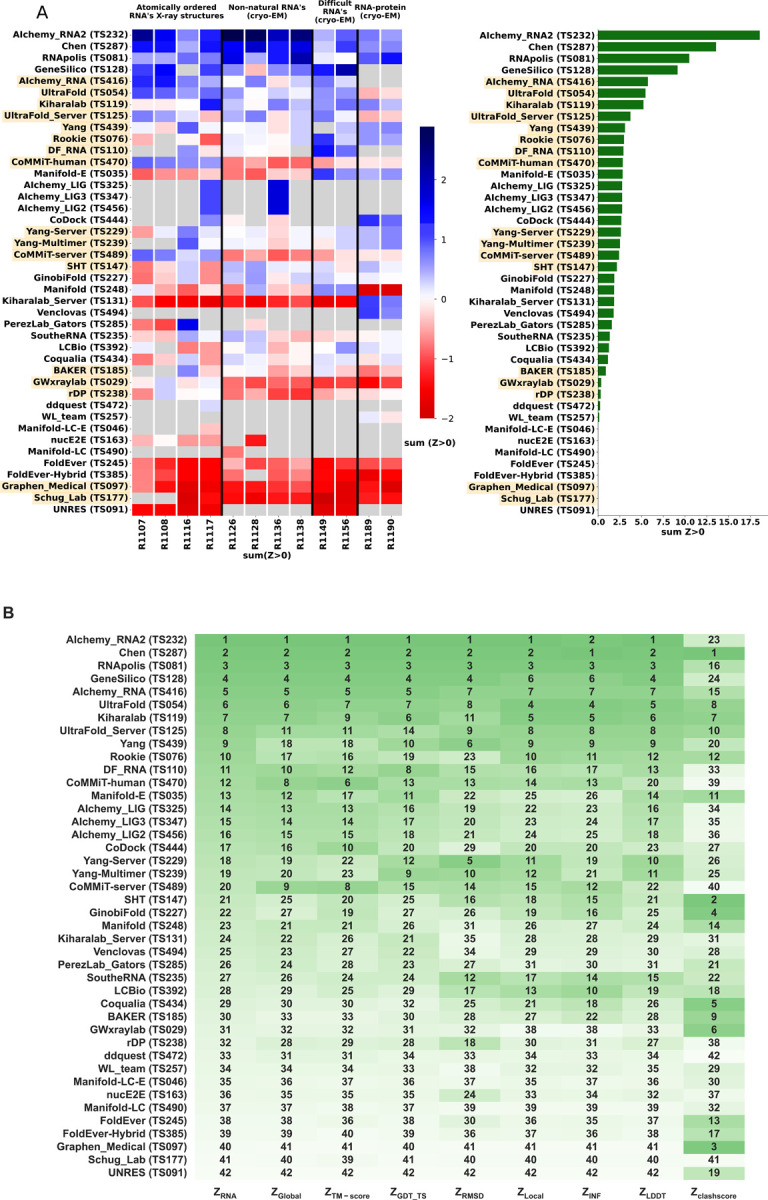
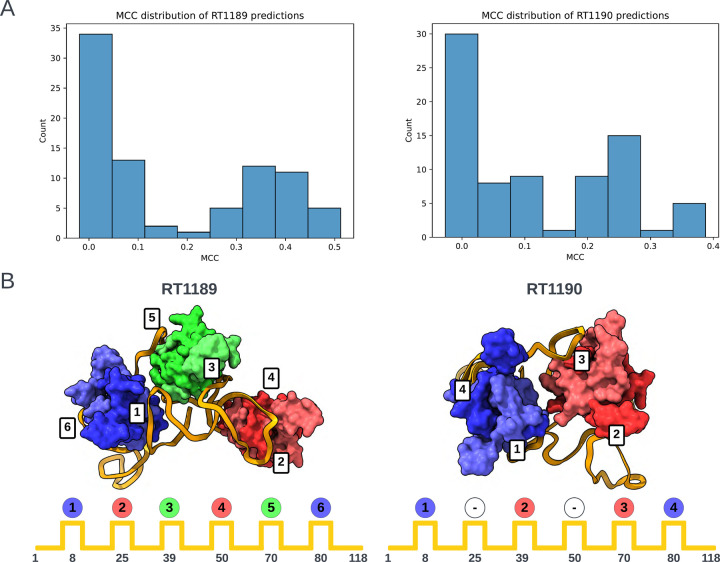
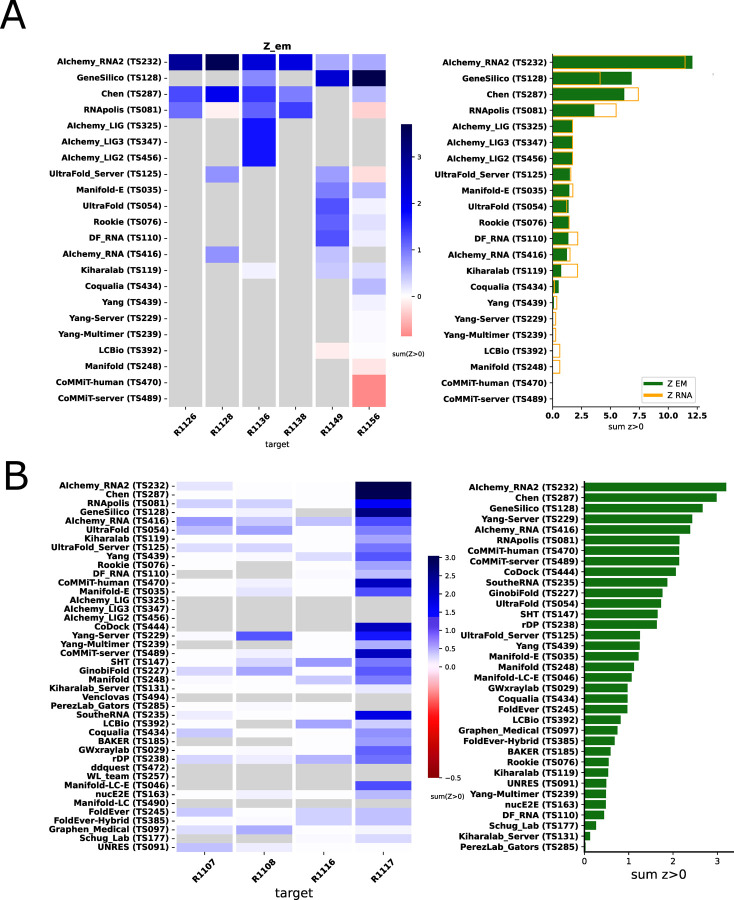
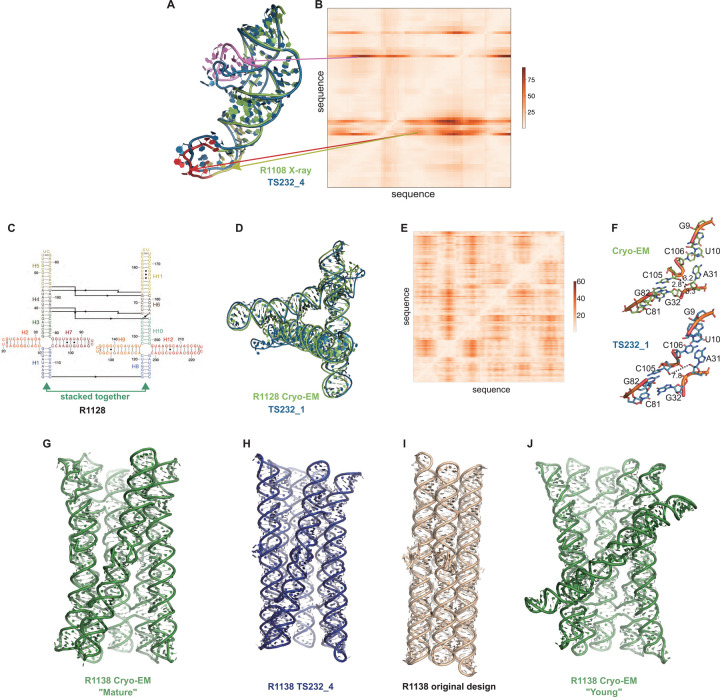
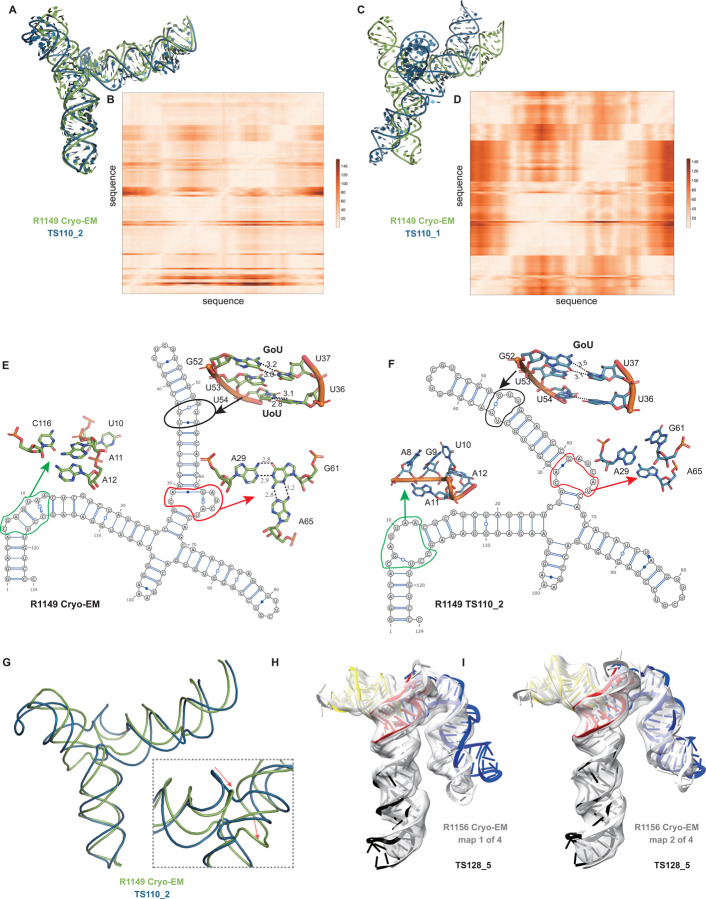
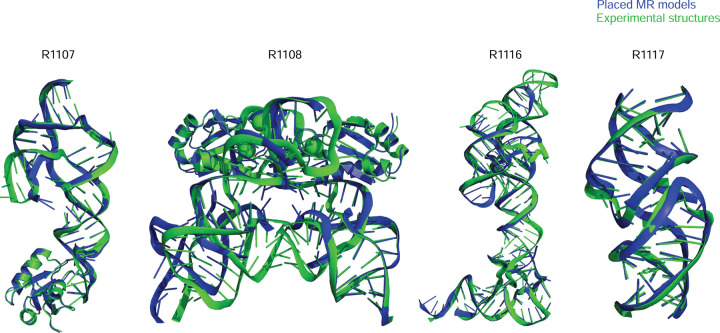
The prediction of RNA three-dimensional structures remains an unsolved problem. Here, we report assessments of RNA structure predictions in CASP15, the first CASP exercise that involved RNA structure modeling. Forty two predictor groups submitted models for at least one of twelve RNA-containing targets. These models were evaluated by the RNA-Puzzles organizers and, separately, by a CASP-recruited team using metrics (GDT, lDDT) and approaches (Z-score rankings) initially developed for assessment of proteins and generalized here for RNA assessment. The two assessments independently ranked the same predictor groups as first (AIchemy_RNA2), second (Chen), and third (RNAPolis and GeneSilico, tied); predictions from deep learning approaches were significantly worse than these top ranked groups, which did not use deep learning. Further analyses based on direct comparison of predicted models to cryogenic electron microscopy (cryo-EM) maps and X-ray diffraction data support these rankings. With the exception of two RNA-protein complexes, models submitted by CASP15 groups correctly predicted the global fold of the RNA targets. Comparisons of CASP15 submissions to designed RNA nanostructures as well as molecular replacement trials highlight the potential utility of current RNA modeling approaches for RNA nanotechnology and structural biology, respectively. Nevertheless, challenges remain in modeling fine details such as non-canonical pairs, in ranking among submitted models, and in prediction of multiple structures resolved by cryo-EM or crystallography.
Conflict of interest statement
Conflict of Interest All authors declare that they have no competing interests.
Figures

Similar articles
-
Assessment of three-dimensional RNA structure prediction in CASP15.Proteins. 2023 Dec;91(12):1747-1770. doi: 10.1002/prot.26602. Epub 2023 Oct 24. Proteins. 2023. PMID: 37876231 Free PMC article.
-
CASP15 cryo-EM protein and RNA targets: Refinement and analysis using experimental maps.Proteins. 2023 Dec;91(12):1935-1951. doi: 10.1002/prot.26644. Proteins. 2023. PMID: 37994556 Free PMC article.
-
Estimation of model accuracy in CASP15 using the ModFOLDdock server.Proteins. 2023 Dec;91(12):1871-1878. doi: 10.1002/prot.26532. Epub 2023 Jun 14. Proteins. 2023. PMID: 37314190 Free PMC article.
-
AI-Driven Deep Learning Techniques in Protein Structure Prediction.Int J Mol Sci. 2024 Aug 1;25(15):8426. doi: 10.3390/ijms25158426. Int J Mol Sci. 2024. PMID: 39125995 Free PMC article. Review.
-
Smart de novo Macromolecular Structure Modeling from Cryo-EM Maps.J Mol Biol. 2023 May 1;435(9):167967. doi: 10.1016/j.jmb.2023.167967. Epub 2023 Jan 18. J Mol Biol. 2023. PMID: 36681181 Review.
References
-
- Holley R. W., Apgar J., Everett G. A., Madison J. T., Marquisee M., Merrill S. H., Penswick J. R. & Zamir A. STRUCTURE OF A RIBONUCLEIC ACID. Science 147, 1462–1465 (1965). - PubMed
-
- Madison J. T., Everett G. A. & Kung H. Nucleotide sequence of a yeast tyrosine transfer RNA. Science 153, 531–534 (1966). - PubMed
-
- Fuller W. & Hodgson A. Conformation of the anticodon loop intRNA. Nature 215, 817–821 (1967). - PubMed
-
- Levitt M. Detailed molecular model for transfer ribonucleic acid. Nature 224, 759–763 (1969). - PubMed
-
- Hingerty B., Brown R. S. & Jack A. Further refinement of the structure of yeast tRNAPhe. J. Mol. Biol. 124, 523–534 (1978). - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources