

This is a preprint.
CD3+ T-cell: CD14+monocyte complexes are dynamic and increased with HIV and glucose intolerance
- PMID: 37162990
- PMCID: PMC10168203
- DOI: 10.1101/2023.04.24.538020
CD3+ T-cell: CD14+monocyte complexes are dynamic and increased with HIV and glucose intolerance
Update in
-
CD3+ T-cell: CD14+ monocyte complexes are dynamic and increased with HIV and glucose intolerance.J Immunol. 2025 Mar 1;214(3):516-531. doi: 10.1093/jimmun/vkae054. J Immunol. 2025. PMID: 40073149 Free PMC article.
Abstract
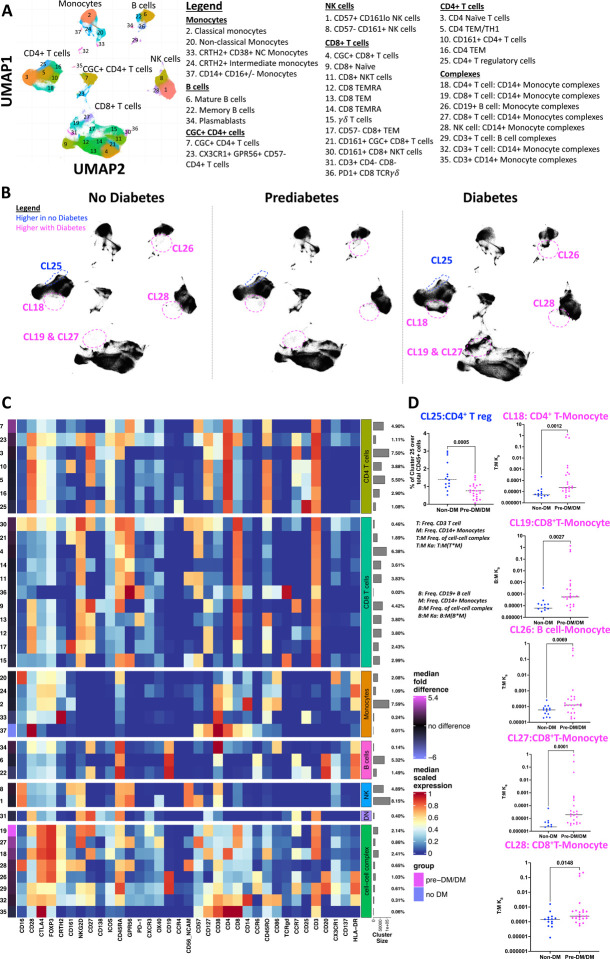
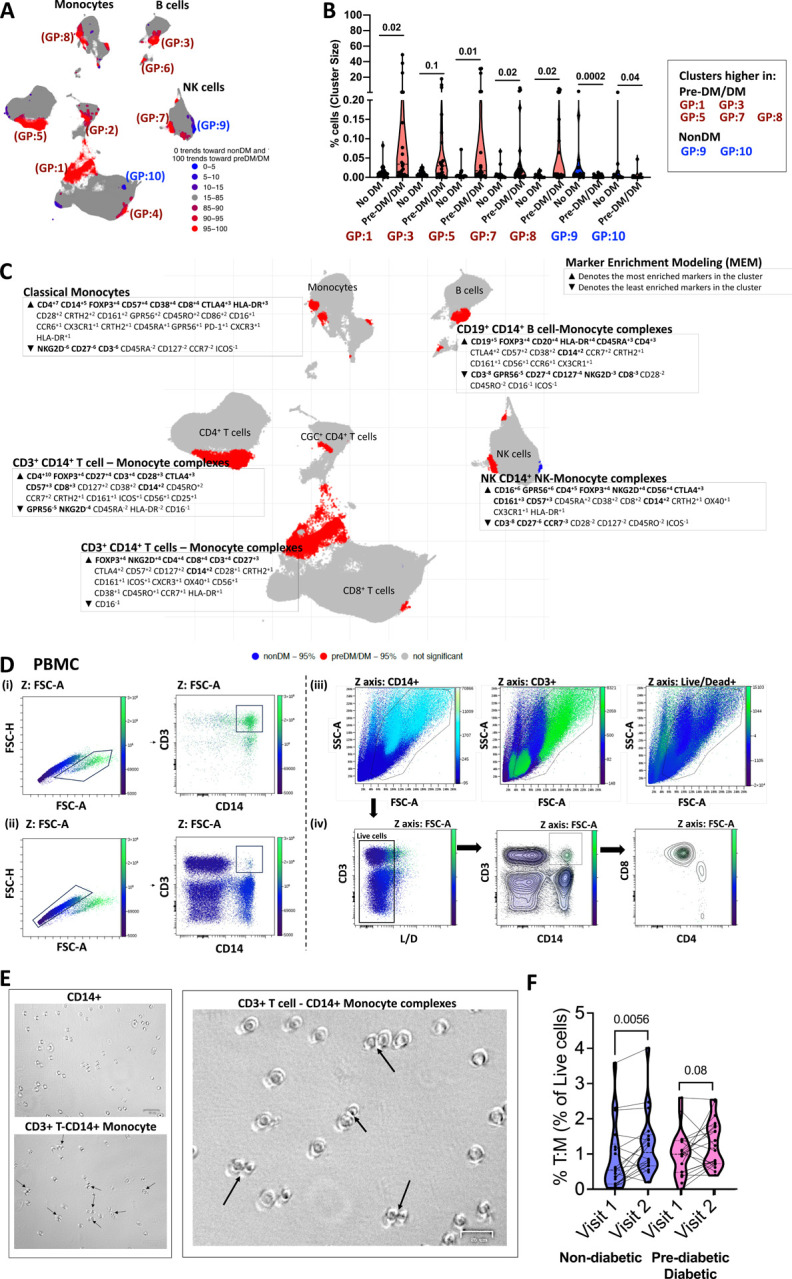
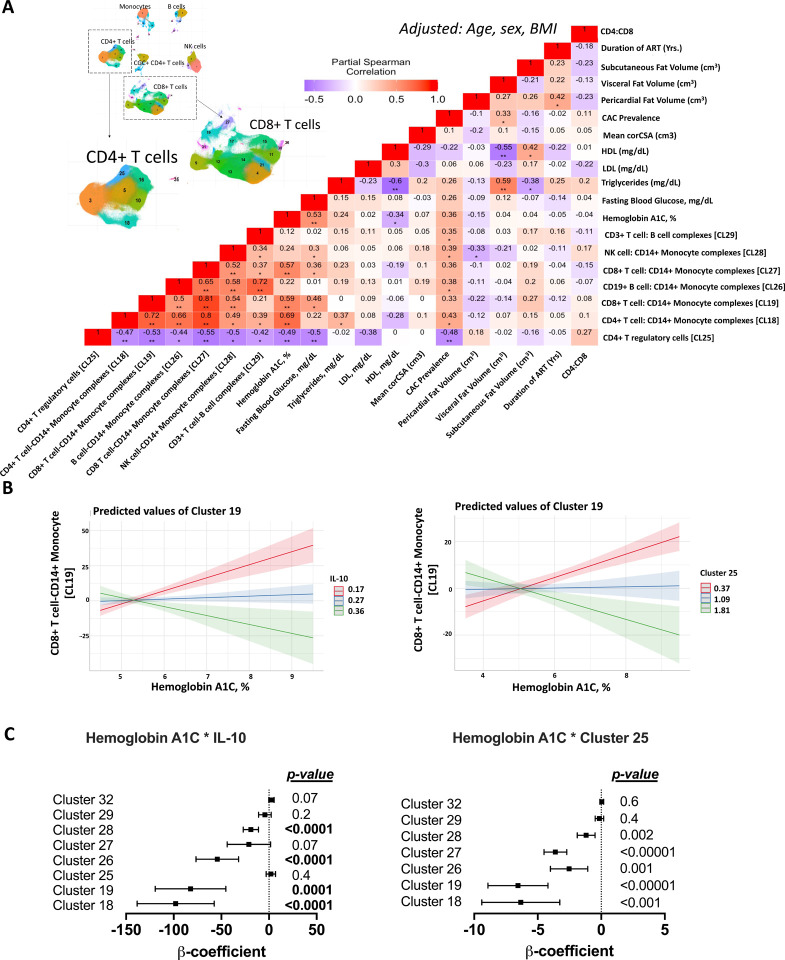
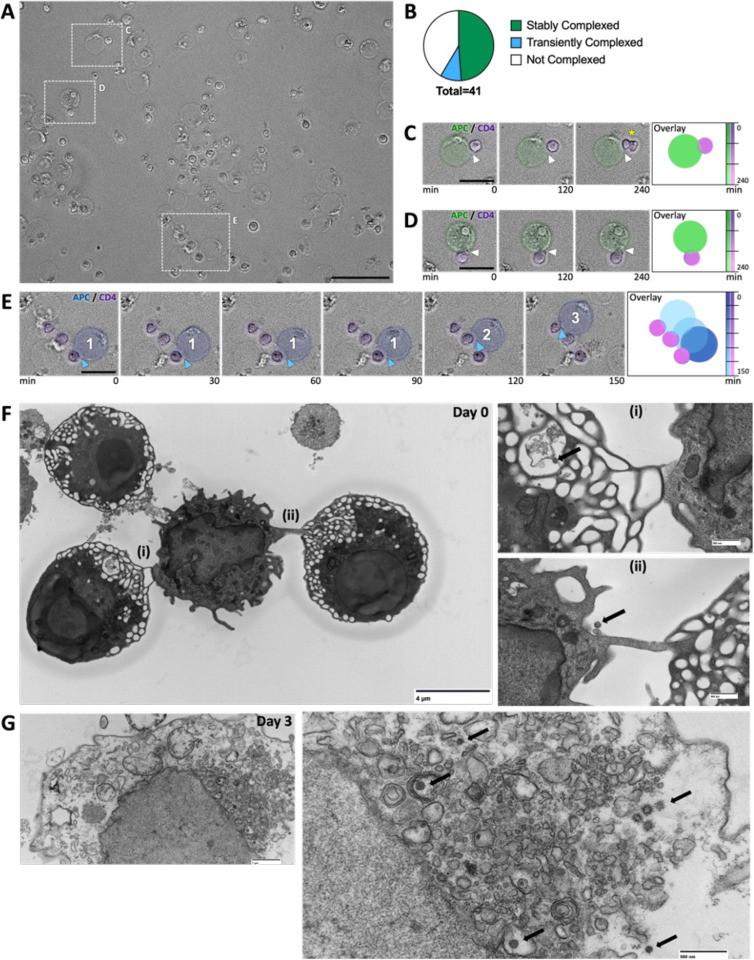
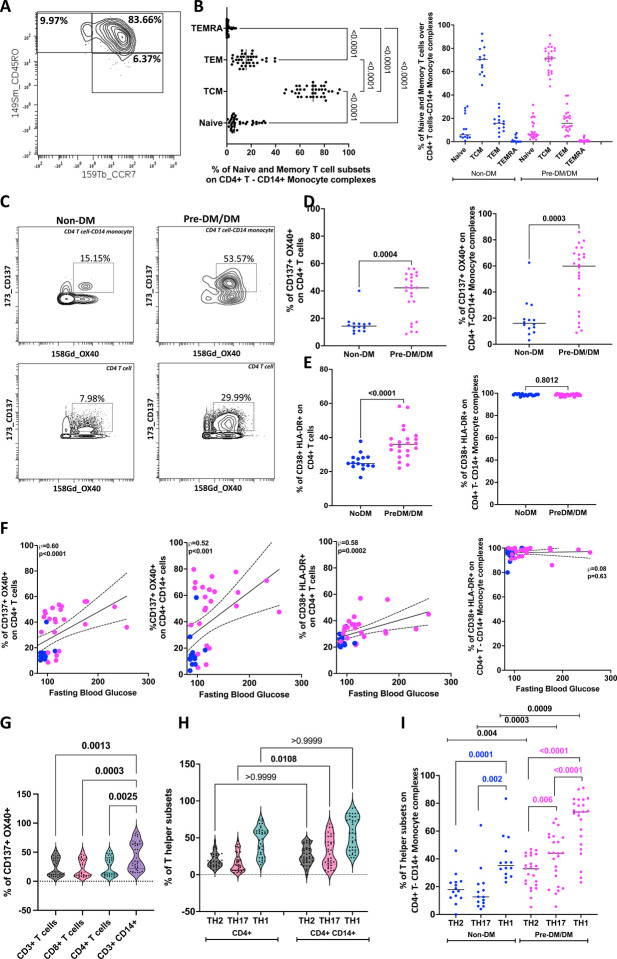
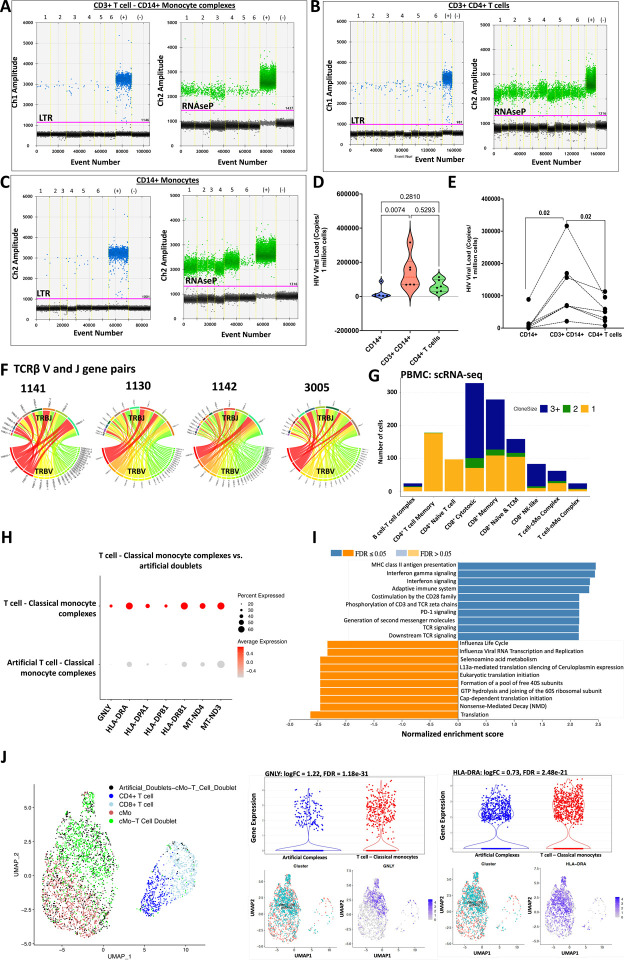
An increased risk of cardiometabolic disease accompanies persistent systemic inflammation. Yet, the innate and adaptive immune system features in persons who develop these conditions remain poorly defined. Doublets, or cell-cell complexes, are routinely eliminated from flow cytometric and other immune phenotyping analyses, which limits our understanding of their relationship to disease states. Using well-characterized clinical cohorts, including participants with controlled HIV as a model for chronic inflammation and increased immune cell interactions, we show that circulating CD14+ monocytes complexed to CD3+ T cells are dynamic, biologically relevant, and increased in individuals with diabetes after adjusting for confounding factors. The complexes form functional immune synapses with increased expression of proinflammatory cytokines and greater glucose utilization. Furthermore, in persons with HIV, the CD3+T-cell: CD14+monocyte complexes had more HIV copies compared to matched CD14+ monocytes or CD4+ T cells alone. Our results demonstrate that circulating CD3+T-cell:CD14+monocyte pairs represent dynamic cellular interactions that may contribute to inflammation and cardiometabolic disease pathogenesis and may originate or be maintained, in part, by chronic viral infections. These findings provide a foundation for future studies investigating mechanisms linking T cellmonocyte cell-cell complexes to developing immune-mediated diseases, including HIV and diabetes.
Keywords: CD3+T-cell:CD14+monocyte complexes; HIV; diabetes; doublets; reservoir.
Conflict of interest statement
Declaration of interests The authors have no competing interests.
Figures
References
-
- Alcaide M. L., Parmigiani A., Pallikkuth S., Roach M., Freguja R., Della Negra M., … Pahwa S. (2013). Immune activation in HIV-infected aging women on antiretrovirals--implications for age-associated comorbidities: a cross-sectional pilot study. PLoS One, 8(5), e63804. doi:10.1371/journal.pone.0063804 - DOI - PMC - PubMed
-
- Arya S M. D., Kemp SE, Jefferis G. (2019). RANN: Fast Nearest Neighbour Search (Wraps ANN Library) Using L2 Metric. R package version 2.6.1. Retrieved from https://github.com/jefferislab/RANN
Publication types
Grants and funding
- K23 HL159351/HL/NHLBI NIH HHS/United States
- KL2 TR002245/TR/NCATS NIH HHS/United States
- UM1 AI164559/AI/NIAID NIH HHS/United States
- IK2 BX004885/BX/BLRD VA/United States
- R01 DK112262/DK/NIDDK NIH HHS/United States
- R25 HL106365/HL/NHLBI NIH HHS/United States
- R25 HL145817/HL/NHLBI NIH HHS/United States
- R01 HL144941/HL/NHLBI NIH HHS/United States
- T32 HL007444/HL/NHLBI NIH HHS/United States
- K23 HL156759/HL/NHLBI NIH HHS/United States
- P30 DK058404/DK/NIDDK NIH HHS/United States
- K12 HL143956/HL/NHLBI NIH HHS/United States
- K08 AR080808/AR/NIAMS NIH HHS/United States
- P30 DK020593/DK/NIDDK NIH HHS/United States
- P30 CA068485/CA/NCI NIH HHS/United States
- P30 AI110527/AI/NIAID NIH HHS/United States
- R03 HL155041/HL/NHLBI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Research Materials