

This is a preprint.
The release of inhibition model reproduces kinetics and plasticity of neurotransmitter release in central synapses
- PMID: 37163032
- PMCID: PMC10168444
- DOI: 10.21203/rs.3.rs-2700789/v1
The release of inhibition model reproduces kinetics and plasticity of neurotransmitter release in central synapses
Update in
-
The release of inhibition model reproduces kinetics and plasticity of neurotransmitter release in central synapses.Commun Biol. 2023 Oct 27;6(1):1091. doi: 10.1038/s42003-023-05445-2. Commun Biol. 2023. PMID: 37891212 Free PMC article.
Abstract
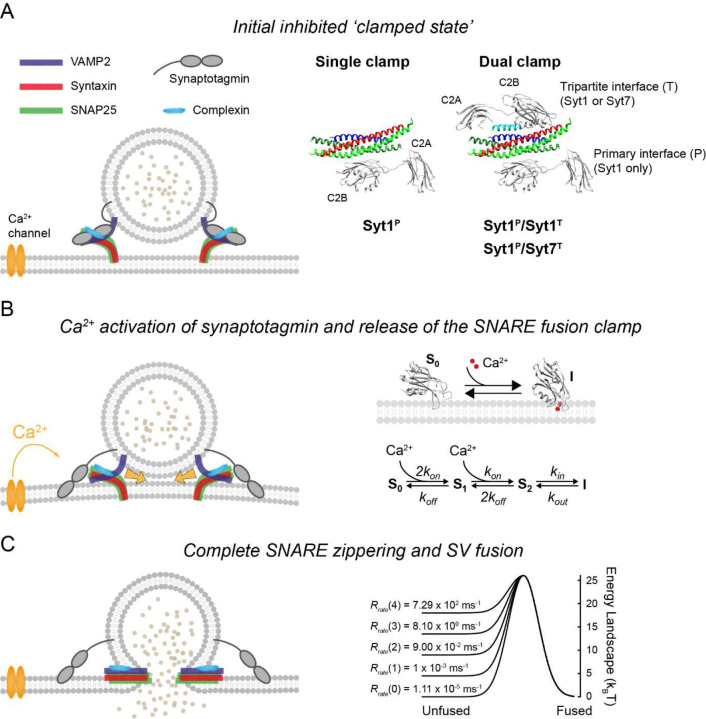
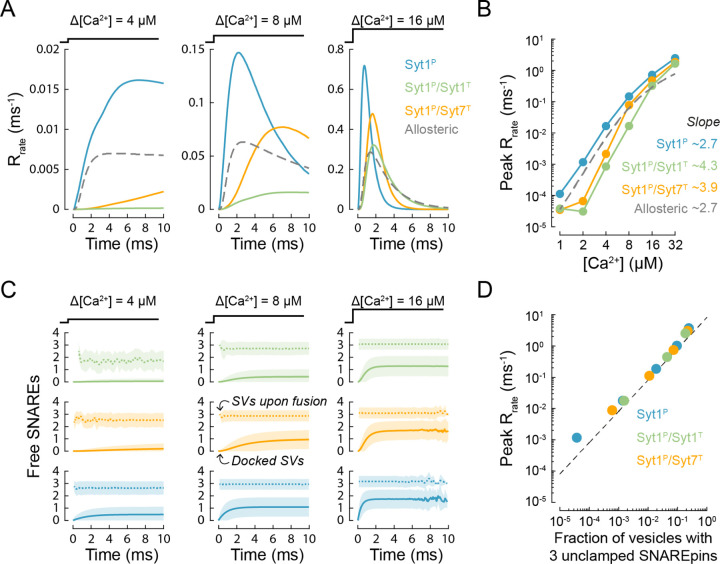
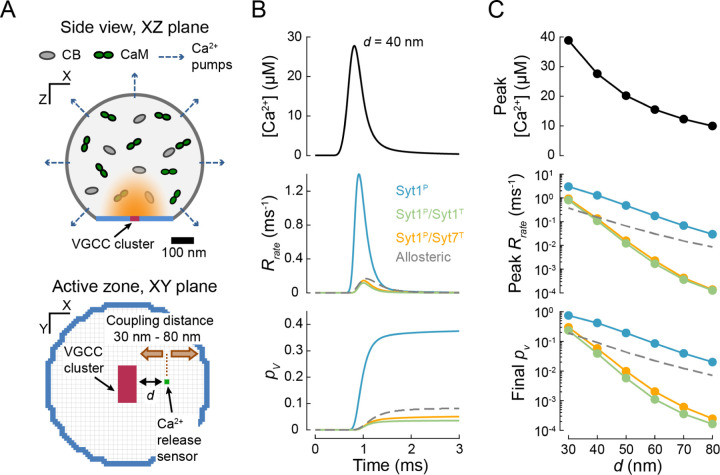
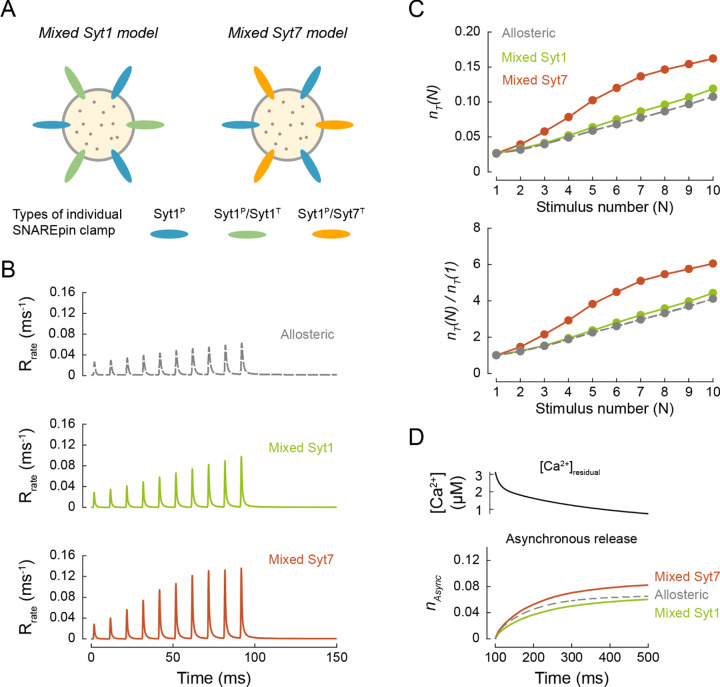
Calcium-evoked release of neurotransmitters from synaptic vesicles (SVs) is catalysed by SNARE proteins. The predominant view is that, at rest, complete assembly of SNARE complexes is inhibited ('clamped') by synaptotagmin and complexin molecules. Calcium binding by synaptotagmins releases this fusion clamp and triggers fast SV exocytosis. However, this model has not been quantitatively tested over physiological timescales. Here we describe an experimentally constrained computational modelling framework to quantitatively assess how the molecular architecture of the fusion clamp affects SV exocytosis. Our results argue that the "release-of-inhibition" model can indeed account for fast calcium-activated SV fusion, and that dual binding of synaptotagmin-1 and synaptotagmin-7 to the same SNARE complex enables synergistic regulation of the kinetics and plasticity of neurotransmitter release. The developed framework provides a powerful and adaptable tool to link the molecular biochemistry of presynaptic proteins to physiological data and efficiently test the plausibility of calcium-activated neurotransmitter release models.
Conflict of interest statement
Competing interests The authors declare no competing interests.
Figures


References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
