

Extracellular vesicles of trypomastigotes of Trypanosoma cruzi induce changes in ubiquitin-related processes, cell-signaling pathways and apoptosis
- PMID: 37165081
- PMCID: PMC10171165
- DOI: 10.1038/s41598-023-34820-6
Extracellular vesicles of trypomastigotes of Trypanosoma cruzi induce changes in ubiquitin-related processes, cell-signaling pathways and apoptosis
Erratum in
-
Author Correction: Extracellular vesicles of trypomastigotes of Trypanosoma cruzi induce changes in ubiquitin-related processes, cell-signaling pathways and apoptosis.Sci Rep. 2023 Jun 6;13(1):9187. doi: 10.1038/s41598-023-35751-y. Sci Rep. 2023. PMID: 37280273 Free PMC article. No abstract available.
Abstract
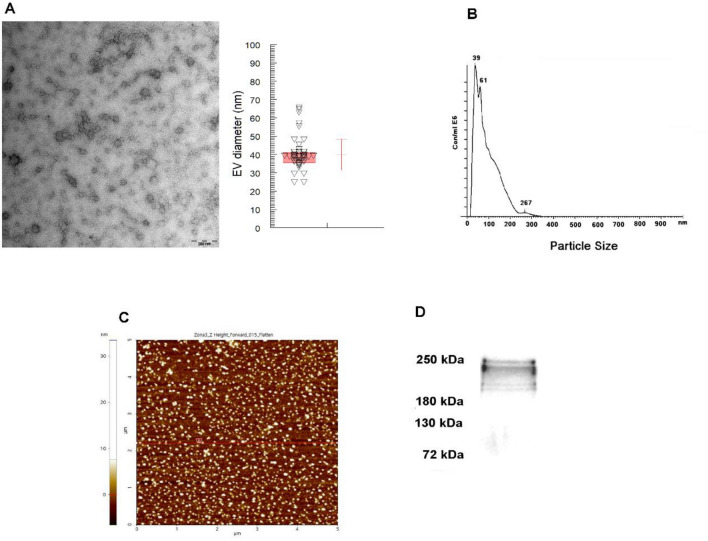
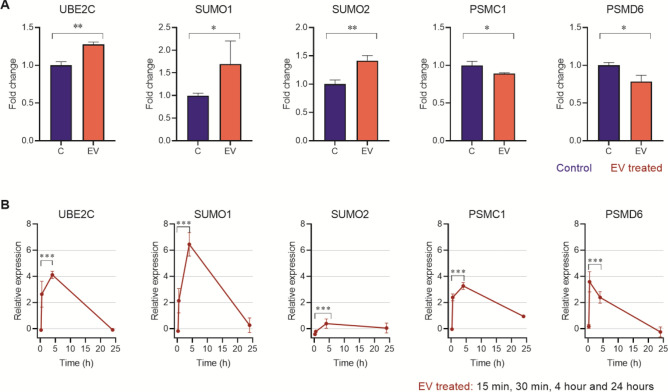
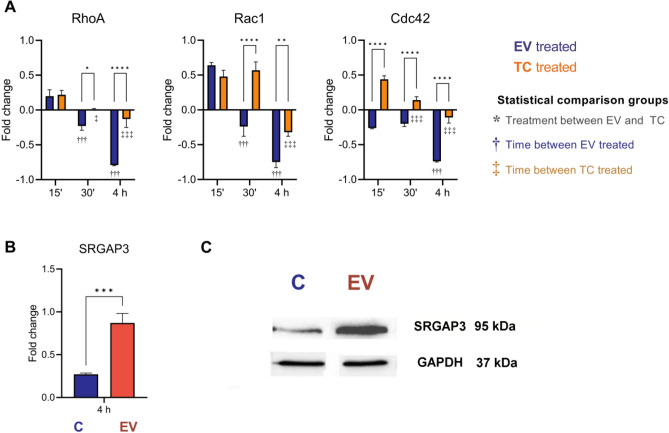
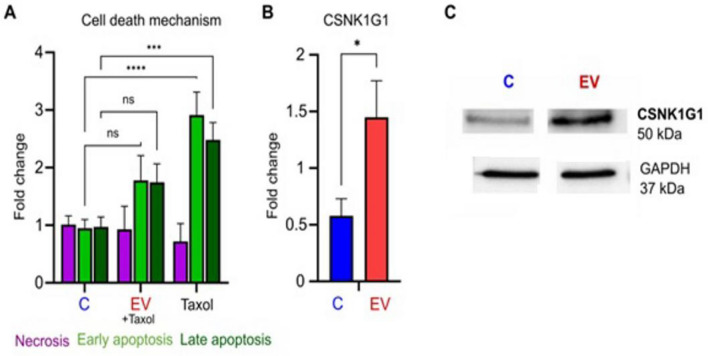
Chagas disease is caused by the protozoan parasite Trypanosoma cruzi. The disease has an acute and a chronic phase in which approximately 30% of the chronic patients suffer from heart disease and/or gastrointestinal symptoms. The pathogenesis of the disease is multifactorial and involves the virulence of the strains, immunological factors and extracellular vesicles (EV) shed by the parasite which participate in cell-cell communication and evasion of the immune response. In this work, we present a transcriptomic analysis of cells stimulated with EV of the trypomastigote stage of T. cruzi. Results after EV-cell incubation revealed 322 differentially expressed genes (168 were upregulated and 154 were downregulated). In this regard, the overexpression of genes related to ubiquitin-related processes (Ube2C, SUMO1 and SUMO2) is highlighted. Moreover, the expression of Rho-GTPases (RhoA, Rac1 and Cdc42) after the interaction was analyzed, revealing a downregulation of the analyzed genes after 4 h of interaction. Finally, a protective role of EV over apoptosis is suggested, as relative values of cells in early and late apoptosis were significantly lower in EV-treated cells, which also showed increased CSNK1G1 expression. These results contribute to a better understanding of the EV-cell interaction and support the role of EV as virulence factors.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures

References
-
- WHO. Chagas disease (American trypanosomiasis) (2021).
-
- WHO Chagas disease in Latin America: An epidemiological update based on 2010 estimates. Wkly. Epidemiol. Rec. 2015;90:33–43. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
