

Single-molecule mechanics and kinetics of cardiac myosin interacting with regulated thin filaments
- PMID: 37165621
- PMCID: PMC10323011
- DOI: 10.1016/j.bpj.2023.05.008
Single-molecule mechanics and kinetics of cardiac myosin interacting with regulated thin filaments
Abstract
The cardiac cycle is a tightly regulated process wherein the heart generates force to pump blood to the body during systole and then relaxes during diastole. Disruption of this finely tuned cycle can lead to a range of diseases including cardiomyopathies and heart failure. Cardiac contraction is driven by the molecular motor myosin, which pulls regulated thin filaments in a calcium-dependent manner. In some muscle and nonmuscle myosins, regulatory proteins on actin tune the kinetics, mechanics, and load dependence of the myosin working stroke; however, it is not well understood whether or how thin-filament regulatory proteins tune the mechanics of the cardiac myosin motor. To address this critical gap in knowledge, we used single-molecule techniques to measure the kinetics and mechanics of the substeps of the cardiac myosin working stroke in the presence and absence of thin filament regulatory proteins. We found that regulatory proteins gate the calcium-dependent interactions between myosin and the thin filament. At physiologically relevant ATP concentrations, cardiac myosin's mechanics and unloaded kinetics are not affected by thin-filament regulatory proteins. We also measured the load-dependent kinetics of cardiac myosin at physiologically relevant ATP concentrations using an isometric optical clamp, and we found that thin-filament regulatory proteins do not affect either the identity or magnitude of myosin's primary load-dependent transition. Interestingly, at low ATP concentrations at both saturating and physiologically relevant subsaturating calcium concentrations, thin-filament regulatory proteins have a small effect on actomyosin dissociation kinetics, suggesting a mechanism beyond simple steric blocking. These results have important implications for the modeling of cardiac physiology and diseases.
Copyright © 2023 Biophysical Society. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
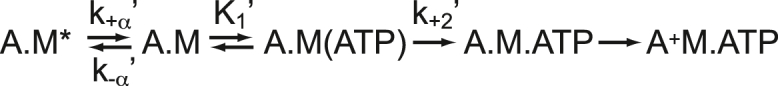





Update of
-
Single Molecule Mechanics and Kinetics of Cardiac Myosin Interacting with Regulated Thin Filaments.bioRxiv [Preprint]. 2023 Jan 10:2023.01.09.522880. doi: 10.1101/2023.01.09.522880. bioRxiv. 2023. Update in: Biophys J. 2023 Jun 20;122(12):2544-2555. doi: 10.1016/j.bpj.2023.05.008. PMID: 36711892 Free PMC article. Updated. Preprint.
Similar articles
-
Single Molecule Mechanics and Kinetics of Cardiac Myosin Interacting with Regulated Thin Filaments.bioRxiv [Preprint]. 2023 Jan 10:2023.01.09.522880. doi: 10.1101/2023.01.09.522880. bioRxiv. 2023. Update in: Biophys J. 2023 Jun 20;122(12):2544-2555. doi: 10.1016/j.bpj.2023.05.008. PMID: 36711892 Free PMC article. Updated. Preprint.
-
Danicamtiv reduces myosin's working stroke but activates the thin filament by accelerating actomyosin attachment.Proc Natl Acad Sci U S A. 2025 Aug 26;122(34):e2515786122. doi: 10.1073/pnas.2515786122. Epub 2025 Aug 20. Proc Natl Acad Sci U S A. 2025. PMID: 40833406
-
Single-myosin crossbridge interactions with actin filaments regulated by troponin-tropomyosin.Proc Natl Acad Sci U S A. 2005 Nov 22;102(47):16990-5. doi: 10.1073/pnas.0506326102. Epub 2005 Nov 15. Proc Natl Acad Sci U S A. 2005. PMID: 16287977 Free PMC article.
-
Regulation of contraction in striated muscle.Physiol Rev. 2000 Apr;80(2):853-924. doi: 10.1152/physrev.2000.80.2.853. Physiol Rev. 2000. PMID: 10747208 Review.
-
Ca(2+)-dependent protein switches in actomyosin based contractile systems.Int J Biochem Cell Biol. 1995 Feb;27(2):97-108. doi: 10.1016/1357-2725(94)00080-u. Int J Biochem Cell Biol. 1995. PMID: 7767791 Review.
Cited by
-
Structural dynamics of the intrinsically disordered linker region of cardiac troponin T.bioRxiv [Preprint]. 2024 Oct 14:2024.05.30.596451. doi: 10.1101/2024.05.30.596451. bioRxiv. 2024. PMID: 38853835 Free PMC article. Preprint.
-
Danicamtiv reduces myosin's working stroke but enhances contraction by activating the thin filament.bioRxiv [Preprint]. 2024 Oct 31:2024.10.09.617269. doi: 10.1101/2024.10.09.617269. bioRxiv. 2024. PMID: 39416013 Free PMC article. Preprint.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
