

Pan-genome survey of the fish pathogen Yersinia ruckeri links accessory- and amplified genes to virulence
- PMID: 37167256
- PMCID: PMC10174560
- DOI: 10.1371/journal.pone.0285257
Pan-genome survey of the fish pathogen Yersinia ruckeri links accessory- and amplified genes to virulence
Abstract
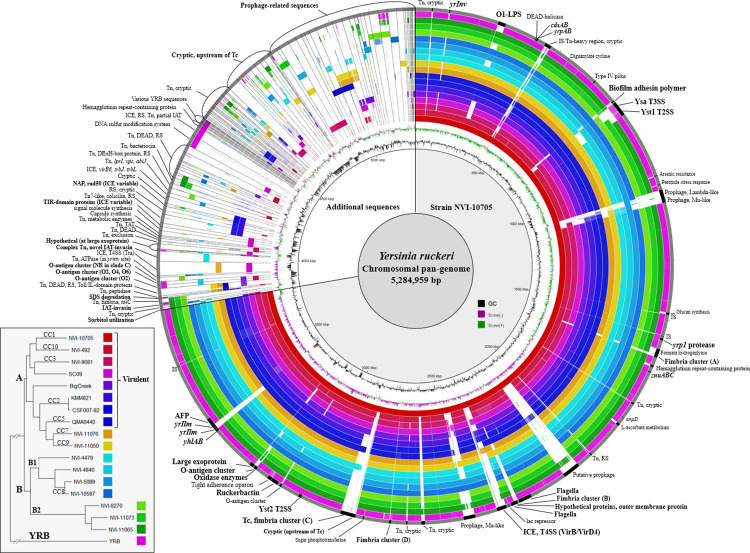
While both virulent and putatively avirulent Yersinia ruckeri strains exist in aquaculture environments, the relationship between the distribution of virulence-associated factors and de facto pathogenicity in fish remains poorly understood. Pan-genome analysis of 18 complete genomes, representing established virulent and putatively avirulent lineages of Y. ruckeri, revealed the presence of a number of accessory genetic determinants. Further investigation of 68 draft genome assemblies revealed that the distribution of certain putative virulence factors correlated well with virulence and host-specificity. The inverse-autotransporter invasin locus yrIlm was, however, the only gene present in all virulent strains, while absent in lineages regarded as avirulent. Strains known to be associated with significant mortalities in salmonid aquaculture display a combination of serotype O1-LPS and yrIlm, with the well-documented highly virulent lineages, represented by MLVA clonal complexes 1 and 2, displaying duplication of the yrIlm locus. Duplication of the yrIlm locus was further found to have evolved over time in clonal complex 1, where some modern, highly virulent isolates display up to three copies.
Copyright: This is an open access article, free of all copyright, and may be freely reproduced, distributed, transmitted, modified, built upon, or otherwise used by anyone for any lawful purpose. The work is made available under the Creative Commons CC0 public domain dedication.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures




References
-
- Sparboe O, Koren C, Håstein T, Poppe TT, Stenwig H. The first isolation of Yersinia ruckeri from farmed Norwegian salmon. Bull Eur Assoc Fish Pathol. 1986;6, 41–42.
-
- Rintamäki P, Valtonen ET, Frerichs GN. Occurrence of Yersinia ruckeri infection in farmed whitefish, Coregonus peled Gmelin and Coregonus muksun Pallas, and Atlantic salmon, Salmo salar L., in northern Finland. J Fish Dis. 1986;9: 137–140. 10.1111/j.1365-2761.1986.tb00993.x - DOI
-
- Costa AA, Leef MJ, Bridle AR, Carson J, Nowak B F. Effect of vaccination against yersiniosis on the relative percent survival, bactericidal and lysozyme response of Atlantic salmon, Salmo salar. Aquaculture. 2011;315, 201–206. 10.1016/j.aquaculture.2011.02.031 - DOI
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
