

Myography of isolated blood vessels: Considerations for experimental design and combination with supplementary techniques
- PMID: 37168231
- PMCID: PMC10165122
- DOI: 10.3389/fphys.2023.1176748
Myography of isolated blood vessels: Considerations for experimental design and combination with supplementary techniques
Abstract
The study of the mechanisms of regulation of vascular tone is an urgent task of modern science, since diseases of the cardiovascular system remain the main cause of reduction in the quality of life and mortality of the population. Myography (isometric and isobaric) of isolated blood vessels is one of the most physiologically relevant approaches to study the function of cells in the vessel wall. On the one hand, cell-cell interactions as well as mechanical stretch of the vessel wall remain preserved in myography studies, in contrast to studies on isolated cells, e.g., cell culture. On the other hand, in vitro studies in isolated vessels allow control of numerous parameters that are difficult to control in vivo. The aim of this review was to 1) discuss the specifics of experimental design and interpretation of data obtained by myography and 2) highlight the importance of the combined use of myography with various complementary techniques necessary for a deep understanding of vascular physiology.
Keywords: artery; endothelium; innervation; intracellular calcium; membrane potential; myography; smooth muscle.
Copyright © 2023 Schubert, Gaynullina, Shvetsova and Tarasova.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
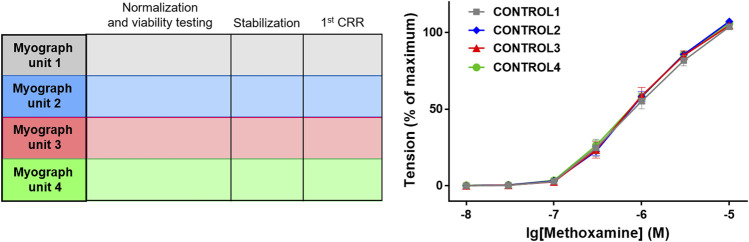
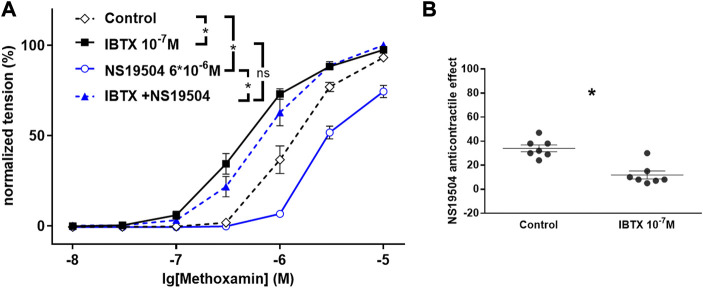
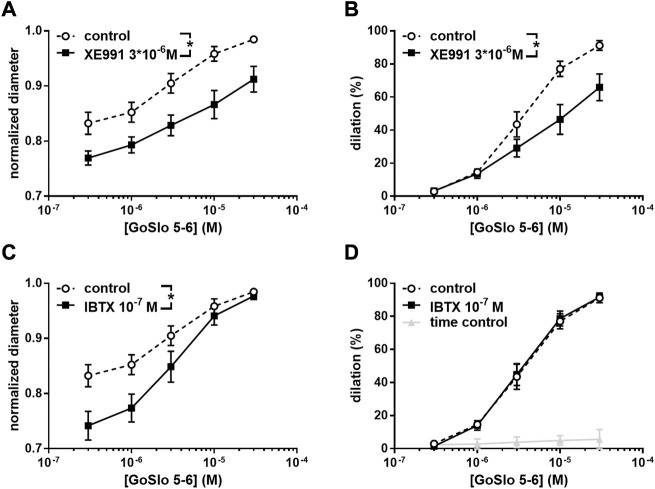
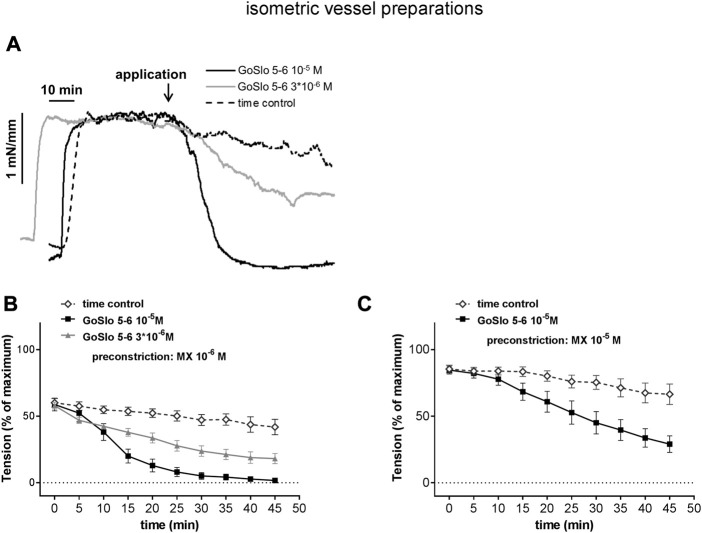
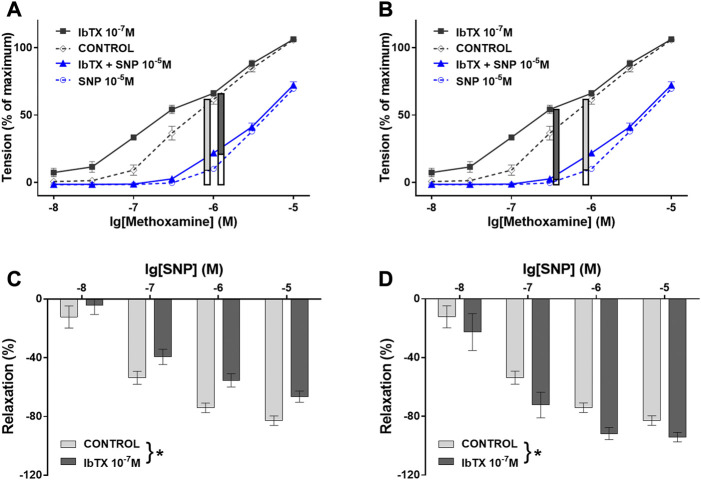
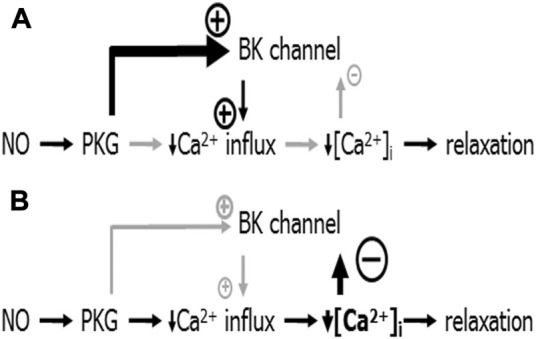

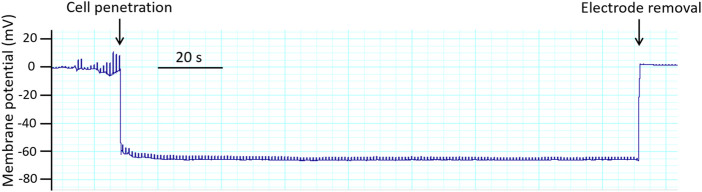
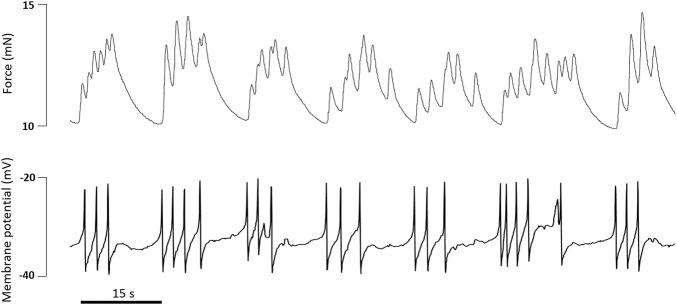
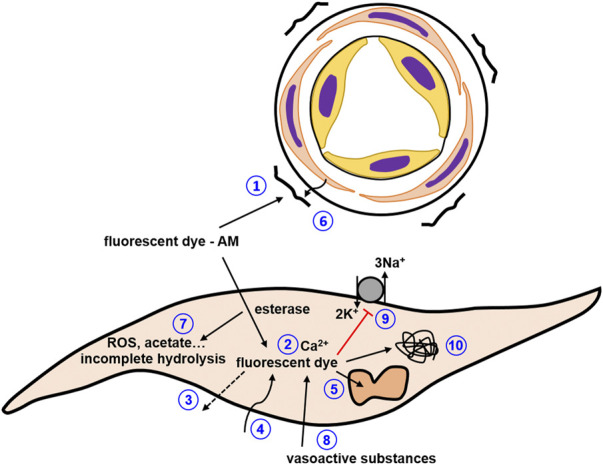
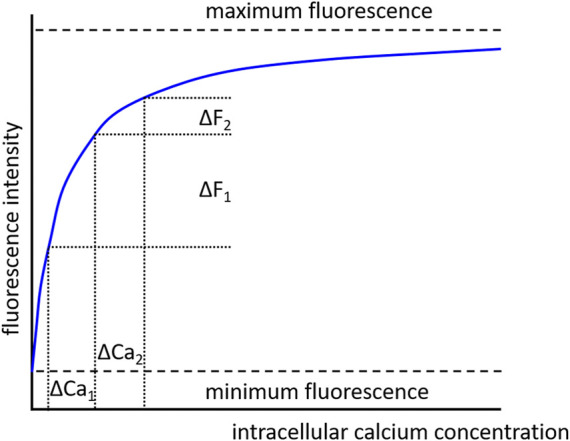
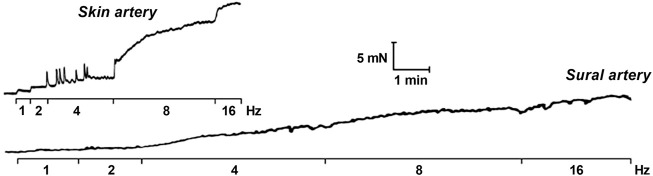
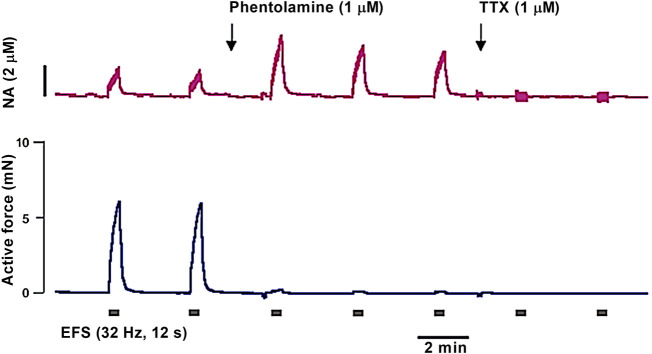
References
Publication types
LinkOut - more resources
Full Text Sources