

An inventory of crosstalk between ubiquitination and other post-translational modifications in orchestrating cellular processes
- PMID: 37168555
- PMCID: PMC10165199
- DOI: 10.1016/j.isci.2023.106276
An inventory of crosstalk between ubiquitination and other post-translational modifications in orchestrating cellular processes
Abstract
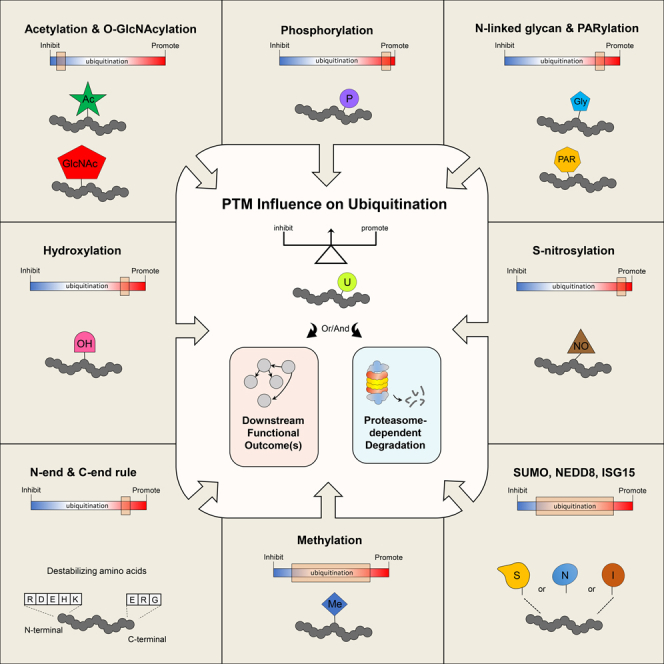
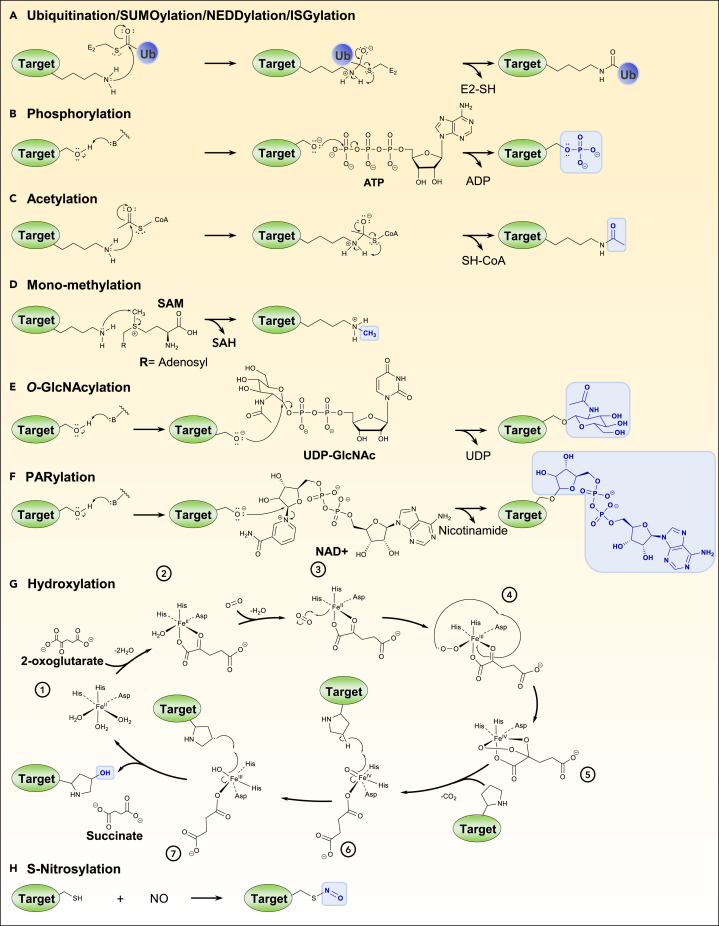
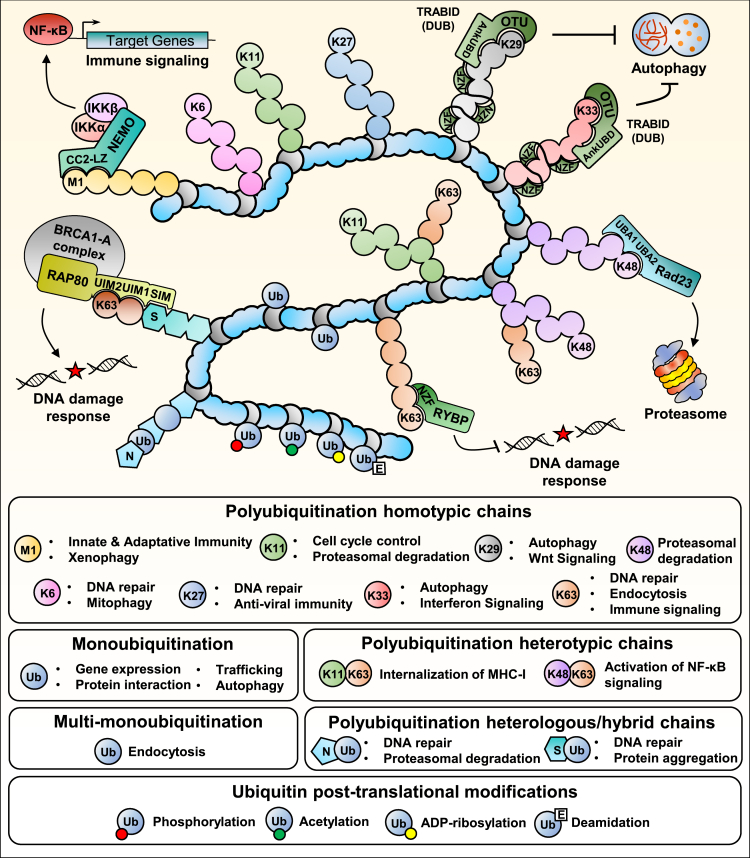
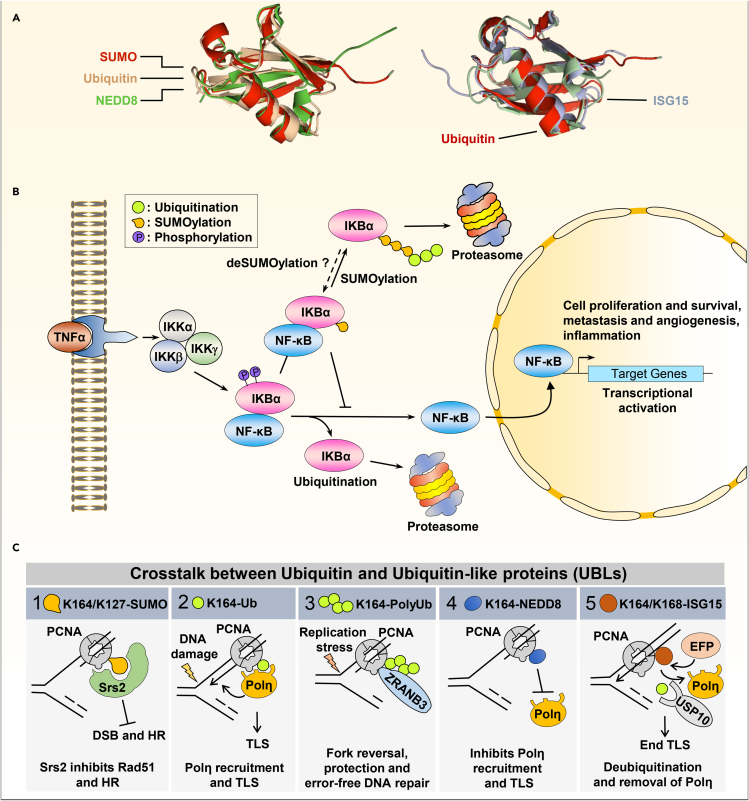
Ubiquitination is an important post-translational modification (PTM) that regulates a large spectrum of cellular processes in eukaryotes. Abnormalities in ubiquitin signaling underlie numerous human pathologies including cancer and neurodegeneration. Much progress has been made during the last three decades in understanding how ubiquitin ligases recognize their substrates and how ubiquitination is orchestrated. Several mechanisms of regulation have evolved to prevent promiscuity including the assembly of ubiquitin ligases in multi-protein complexes with dedicated subunits and specific post-translational modifications of these enzymes and their co-factors. Here, we outline another layer of complexity involving the coordinated access of E3 ligases to substrates. We provide an extensive inventory of ubiquitination crosstalk with multiple PTMs including SUMOylation, phosphorylation, methylation, acetylation, hydroxylation, prolyl isomerization, PARylation, and O-GlcNAcylation. We discuss molecular mechanisms by which PTMs orchestrate ubiquitination, thus increasing its specificity as well as its crosstalk with other signaling pathways to ensure cell homeostasis.
Keywords: Biochemistry; Biological sciences; Cell biology; Molecular biology.
© 2023 The Authors.
Conflict of interest statement
The authors declare no competing interests with the content of this article.
Figures
References
Publication types
LinkOut - more resources
Full Text Sources
Miscellaneous
