

GAP43-dependent mitochondria transfer from astrocytes enhances glioblastoma tumorigenicity
- PMID: 37169842
- PMCID: PMC10212766
- DOI: 10.1038/s43018-023-00556-5
GAP43-dependent mitochondria transfer from astrocytes enhances glioblastoma tumorigenicity
Abstract
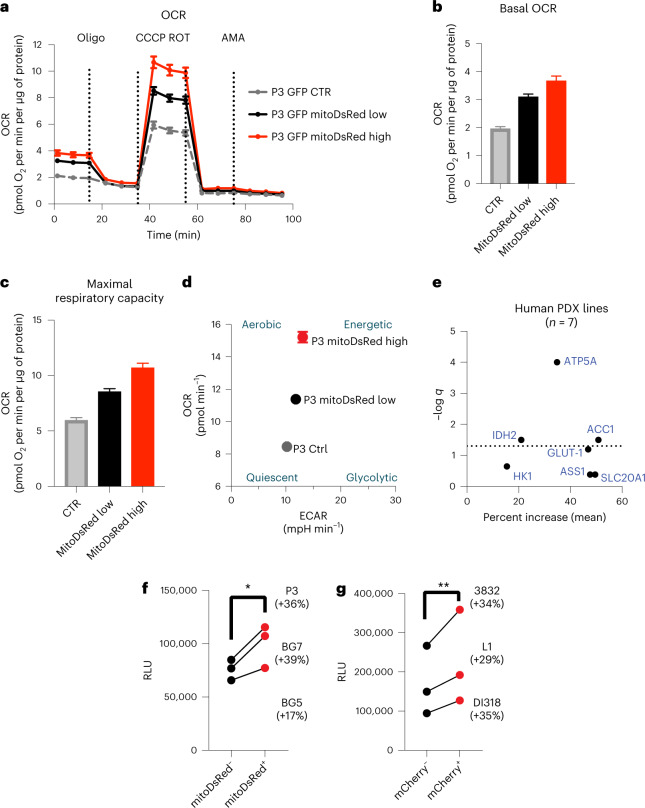
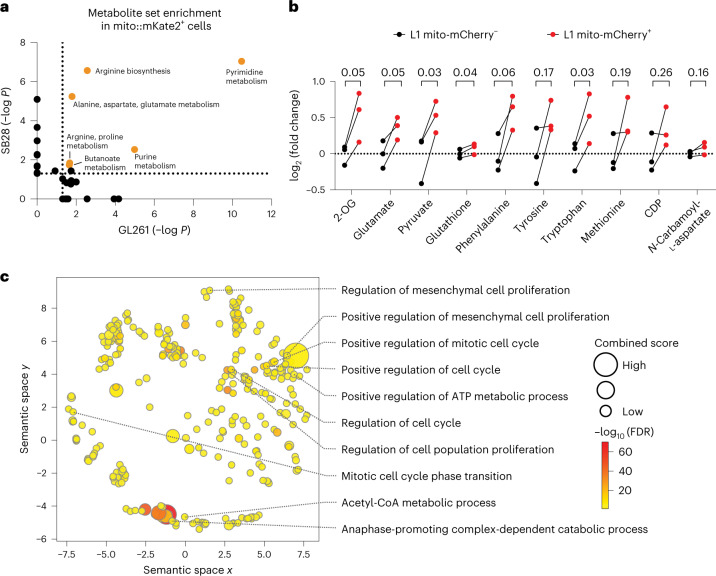
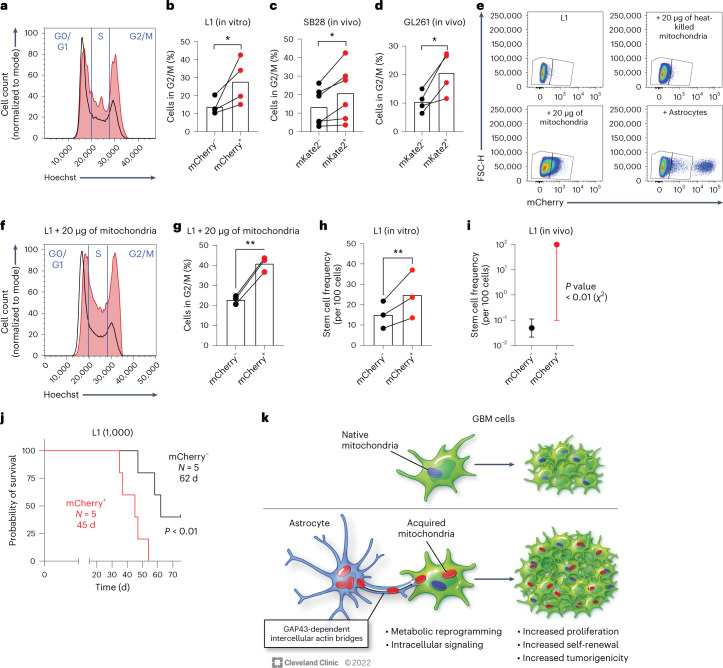
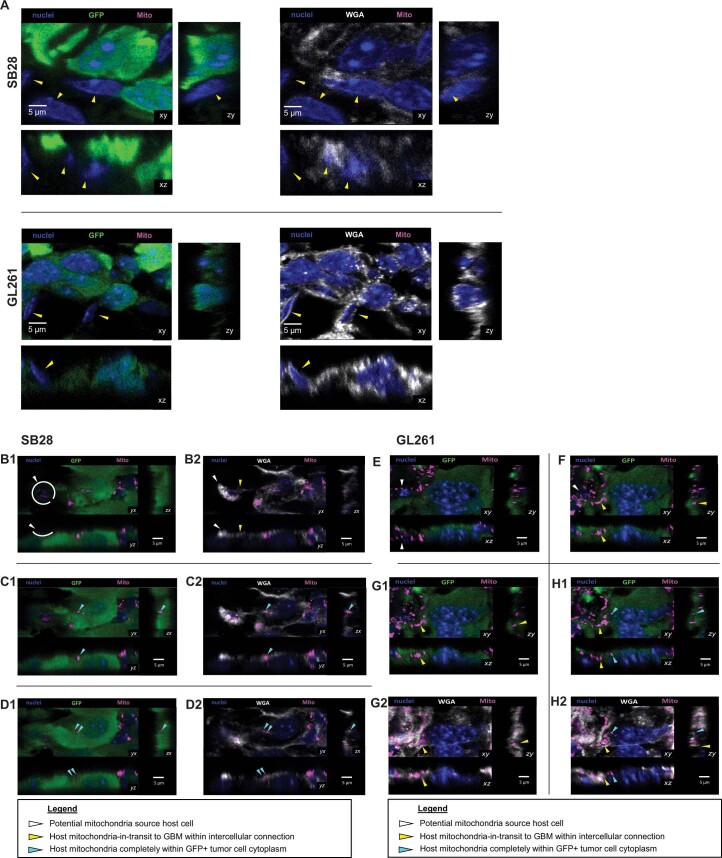
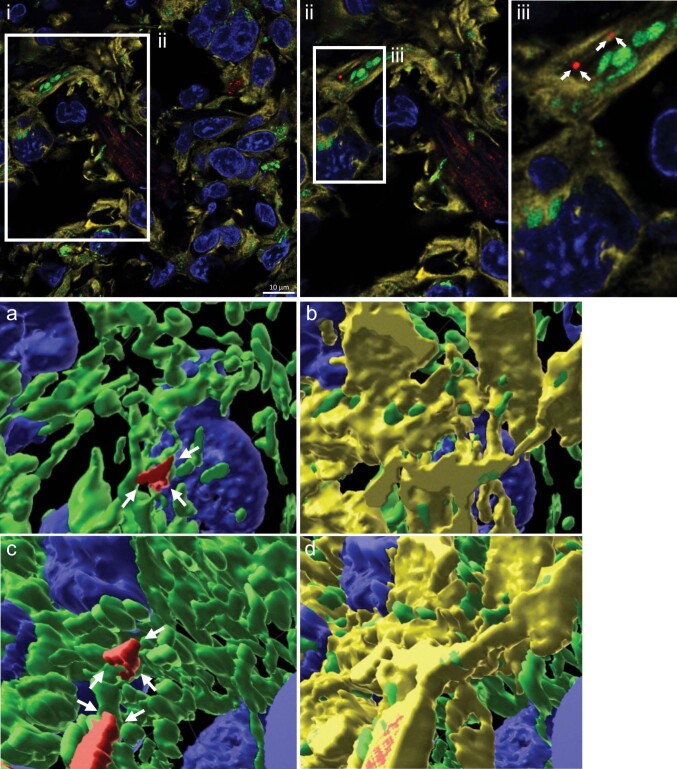
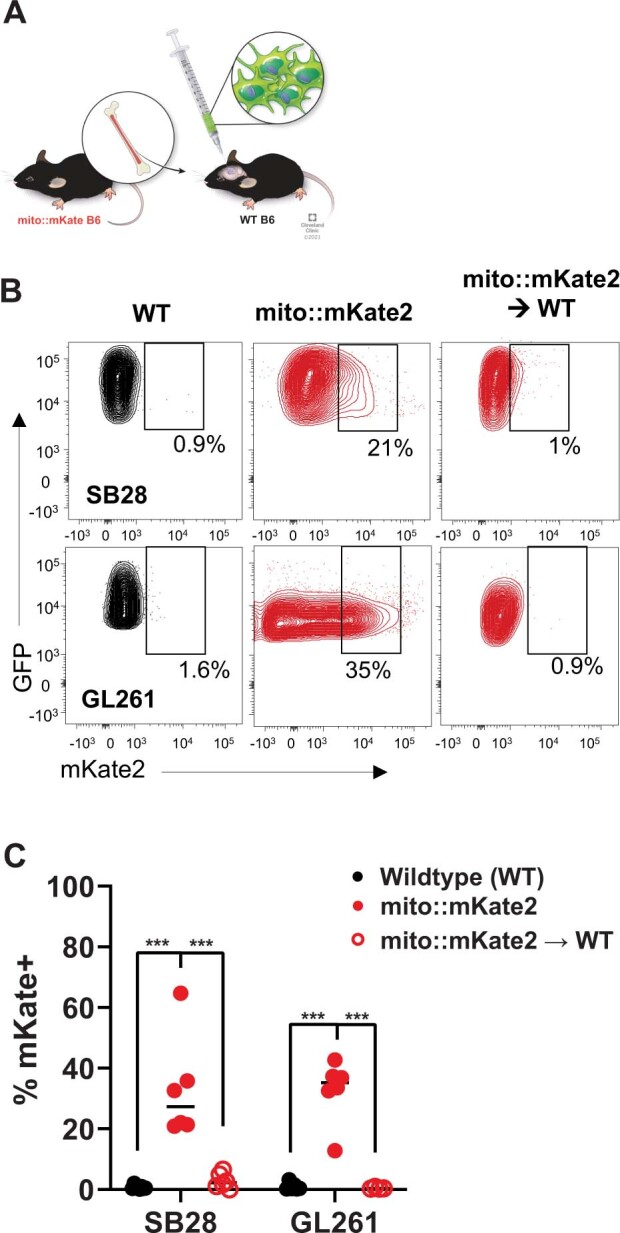
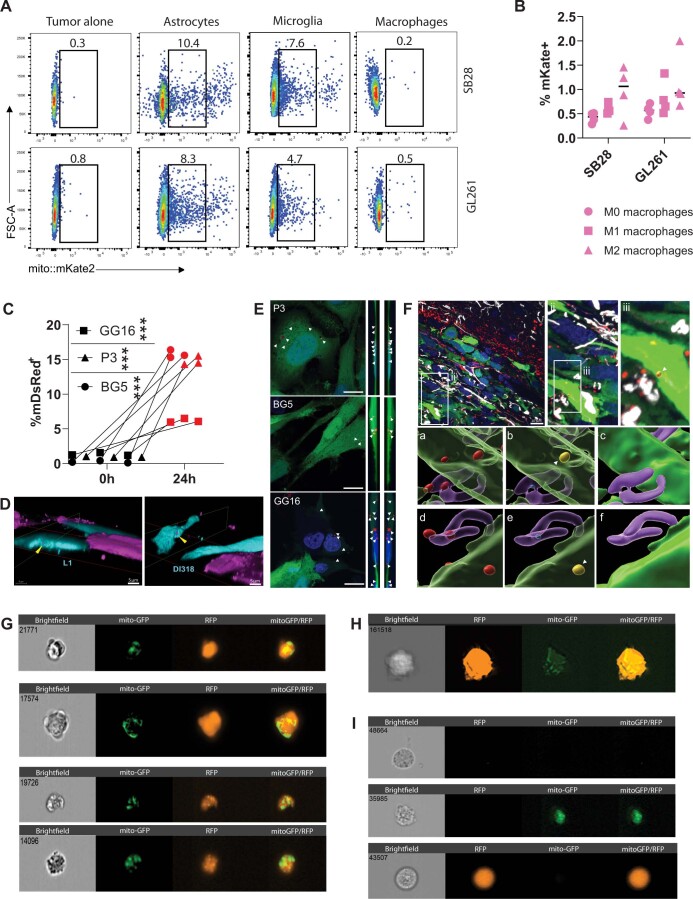
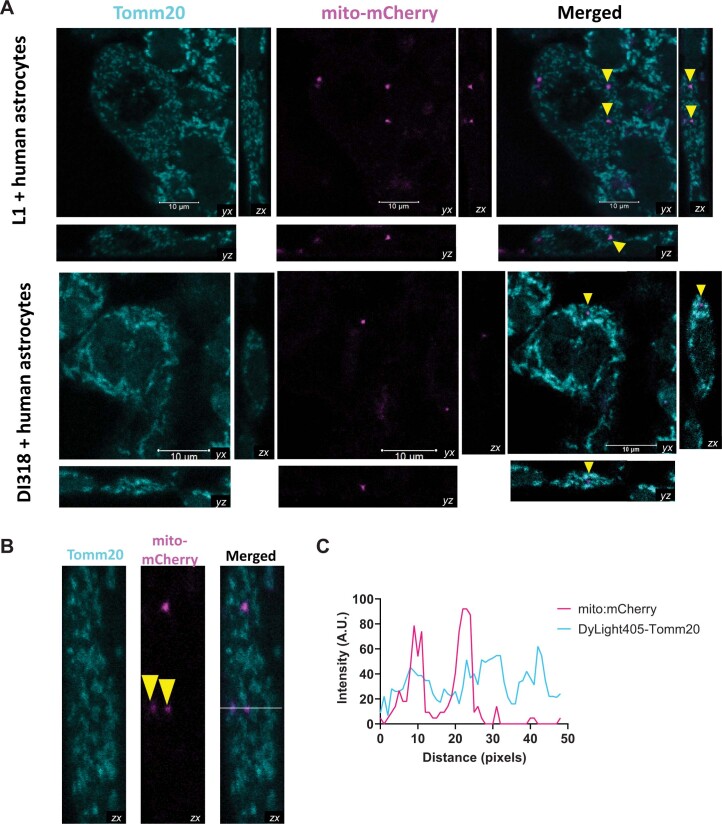
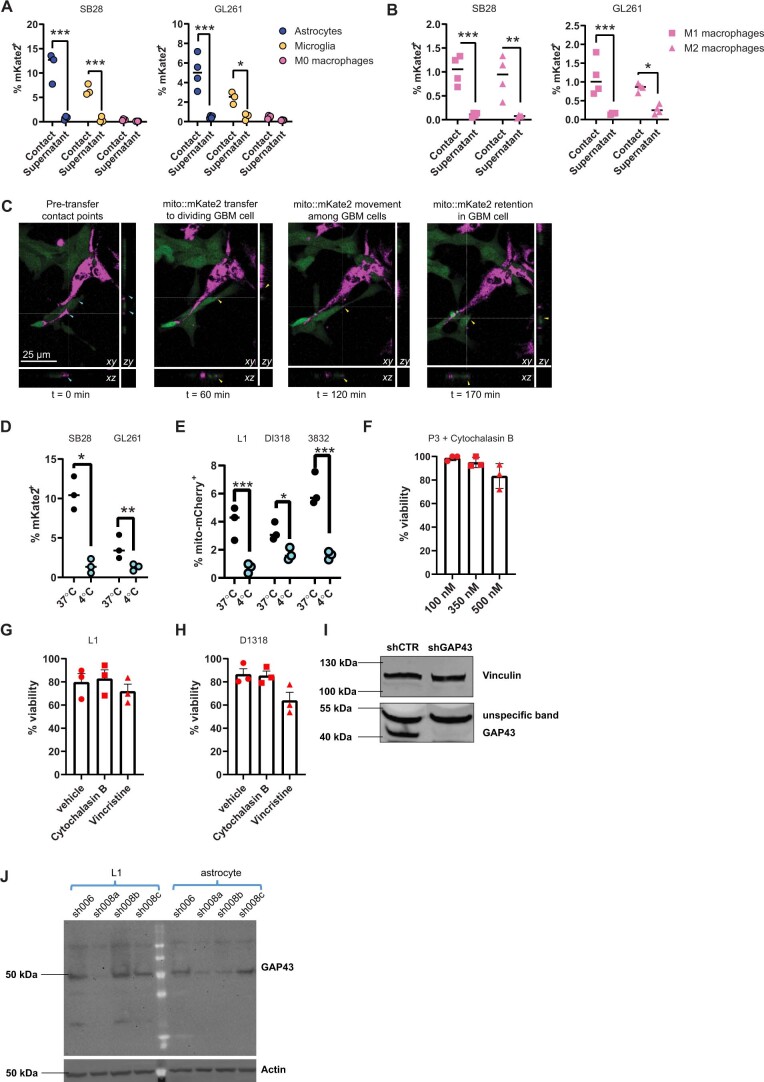
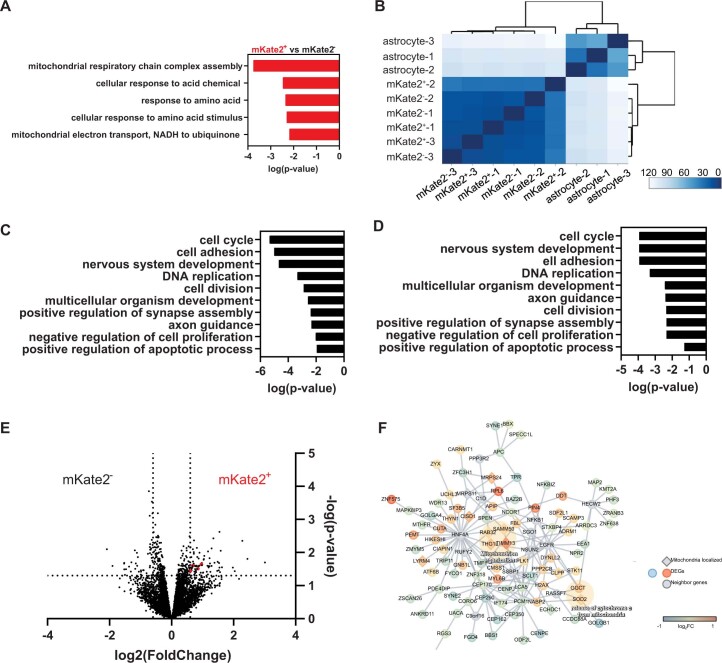
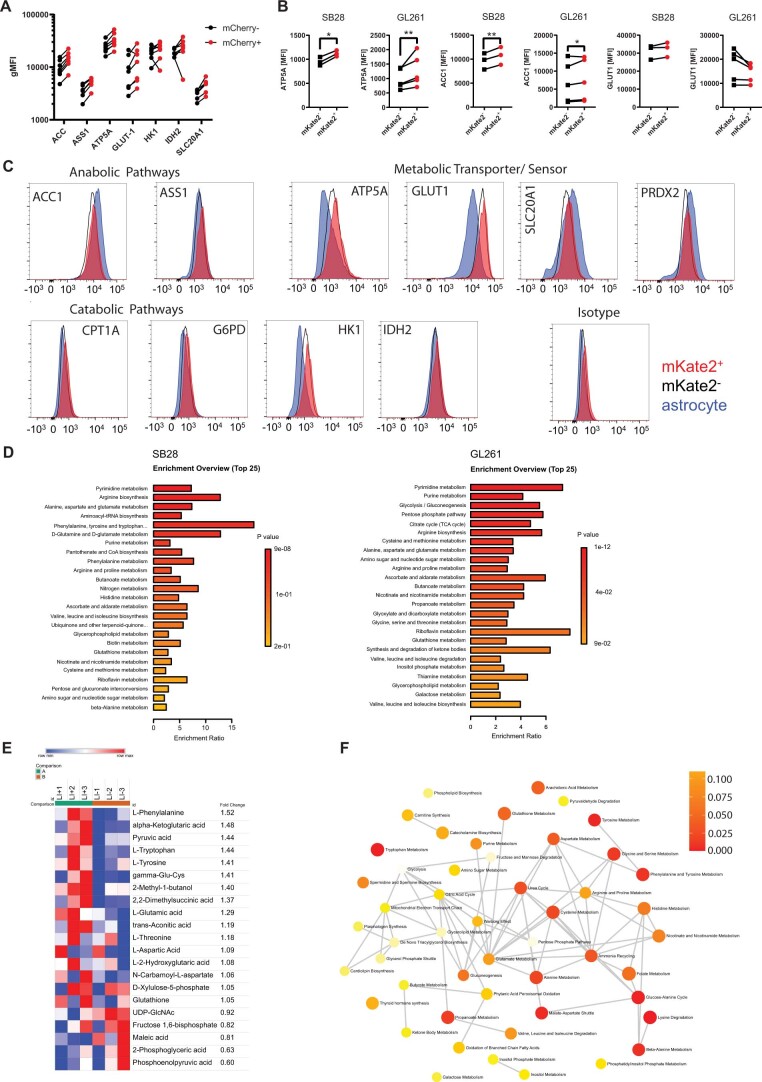
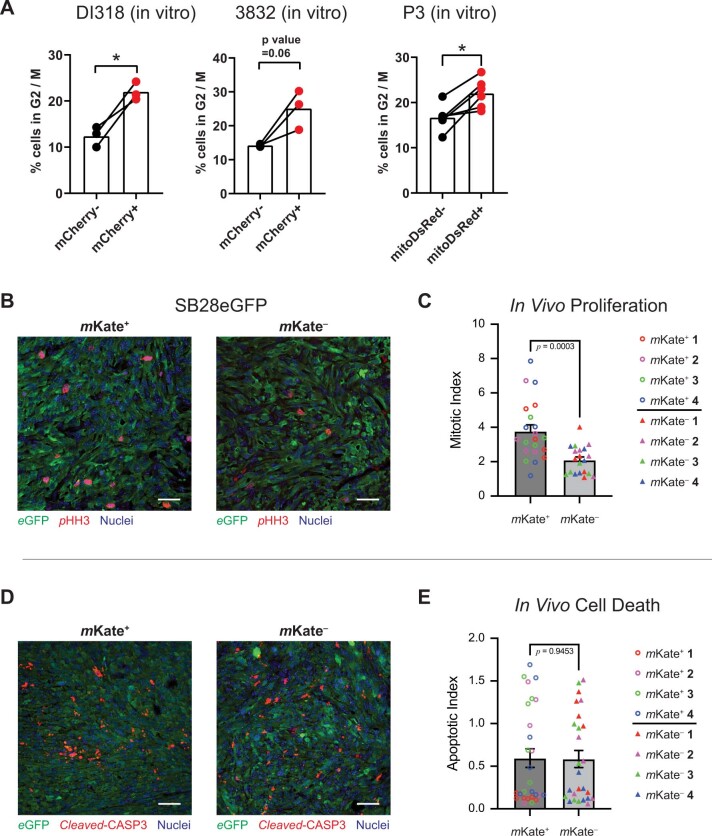
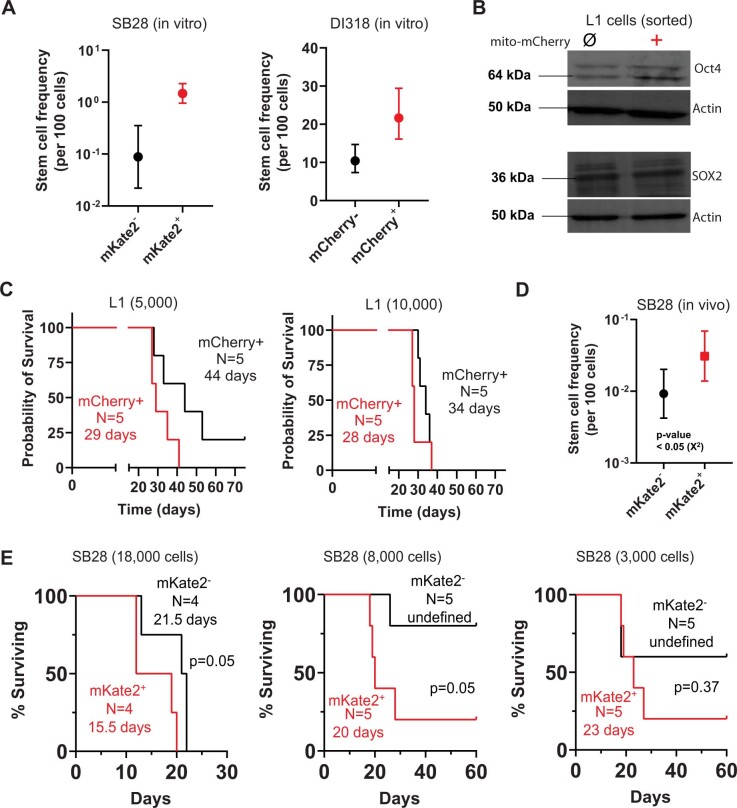
The transfer of intact mitochondria between heterogeneous cell types has been confirmed in various settings, including cancer. However, the functional implications of mitochondria transfer on tumor biology are poorly understood. Here we show that mitochondria transfer is a prevalent phenomenon in glioblastoma (GBM), the most frequent and malignant primary brain tumor. We identified horizontal mitochondria transfer from astrocytes as a mechanism that enhances tumorigenesis in GBM. This transfer is dependent on network-forming intercellular connections between GBM cells and astrocytes, which are facilitated by growth-associated protein 43 (GAP43), a protein involved in neuron axon regeneration and astrocyte reactivity. The acquisition of astrocyte mitochondria drives an increase in mitochondrial respiration and upregulation of metabolic pathways linked to proliferation and tumorigenicity. Functionally, uptake of astrocyte mitochondria promotes cell cycle progression to proliferative G2/M phases and enhances self-renewal and tumorigenicity of GBM. Collectively, our findings reveal a host-tumor interaction that drives proliferation and self-renewal of cancer cells, providing opportunities for therapeutic development.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
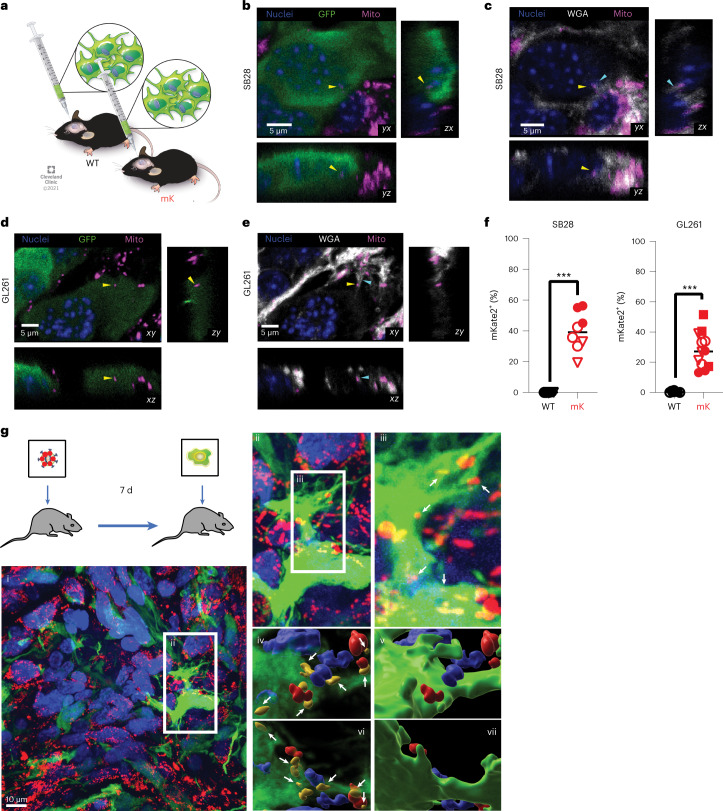
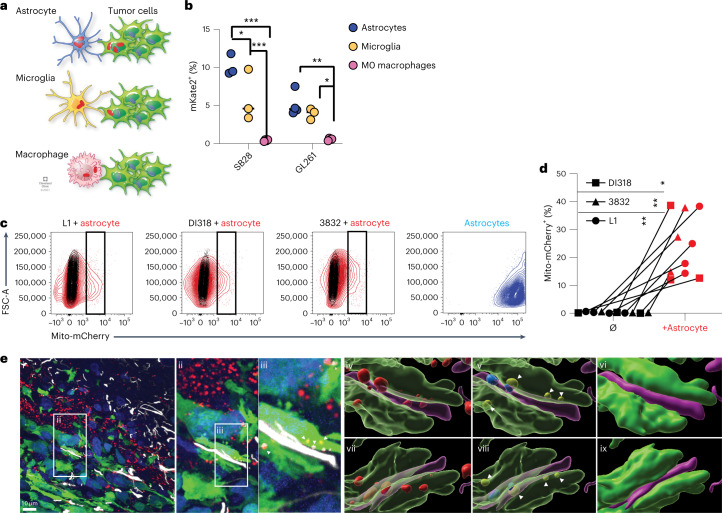
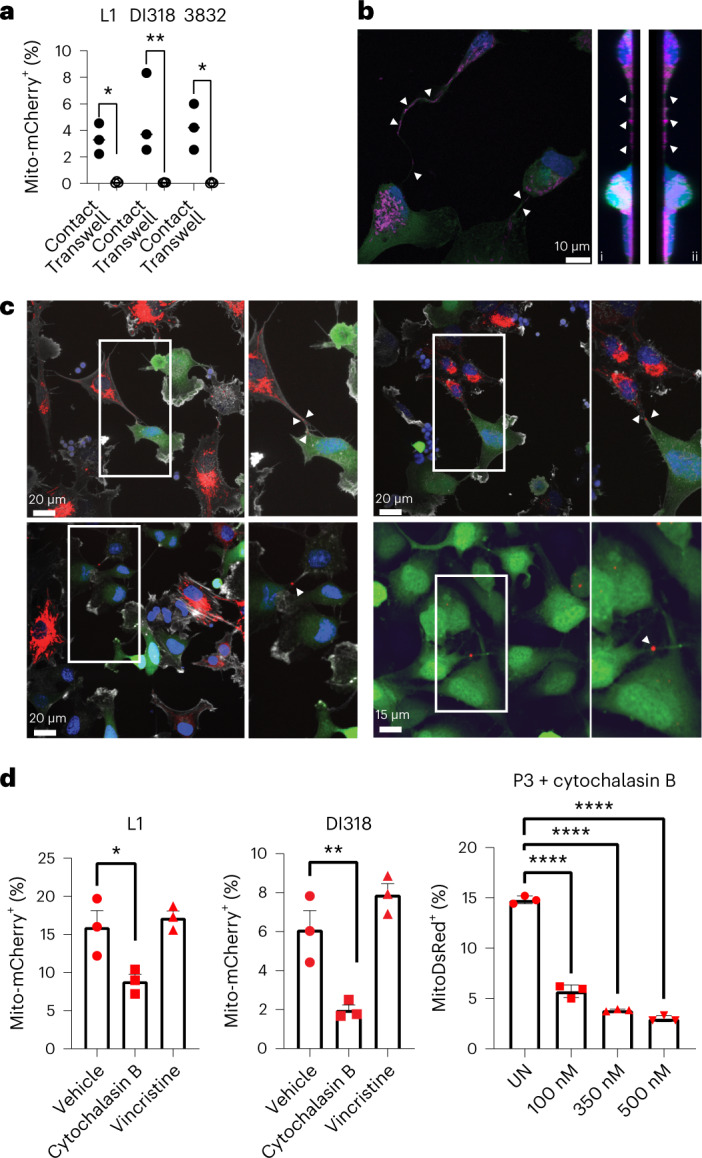
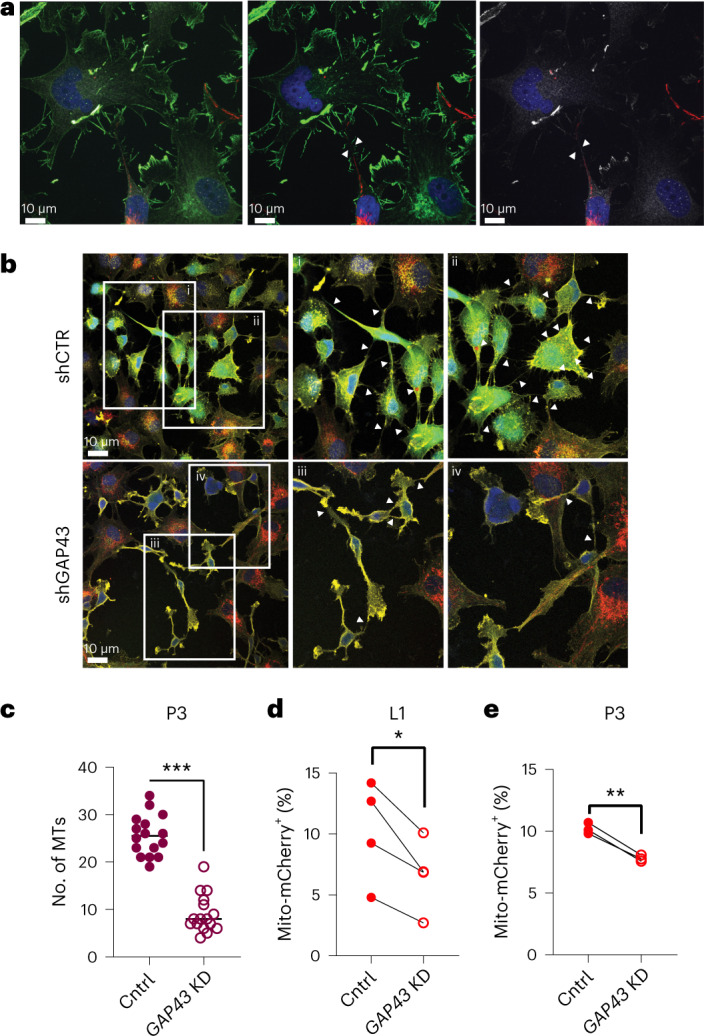
Comment in
-
Mind the GAP(43) for mitochondria transfer to glioblastomas.Nat Cancer. 2023 May;4(5):588-589. doi: 10.1038/s43018-023-00564-5. Nat Cancer. 2023. PMID: 37169844 No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
