

Aerobic bacteria produce nitric oxide via denitrification and promote algal population collapse
- PMID: 37173383
- PMCID: PMC10356946
- DOI: 10.1038/s41396-023-01427-8
Aerobic bacteria produce nitric oxide via denitrification and promote algal population collapse
Abstract
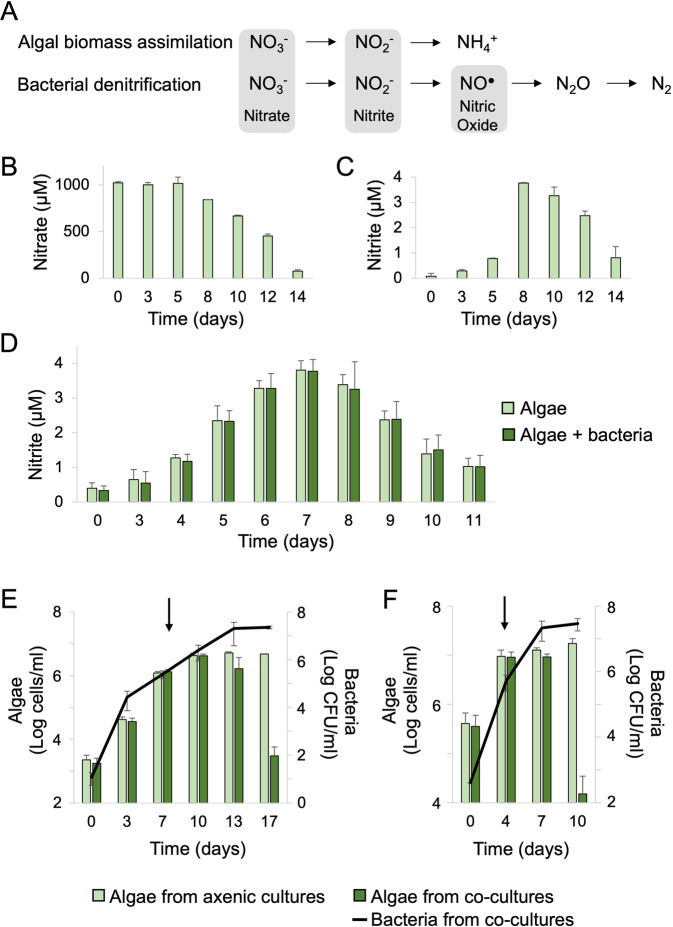
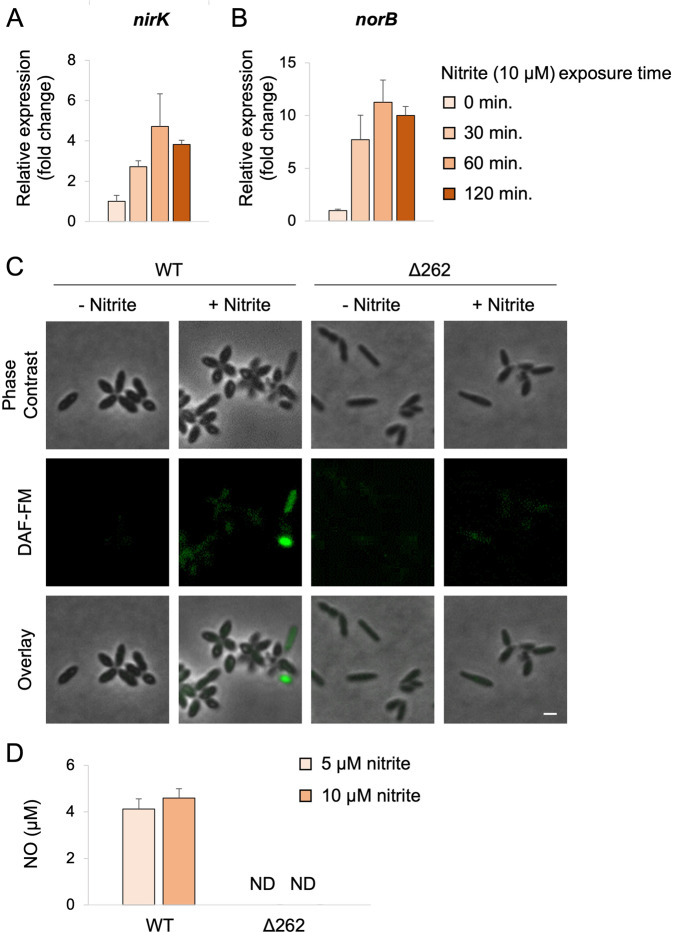
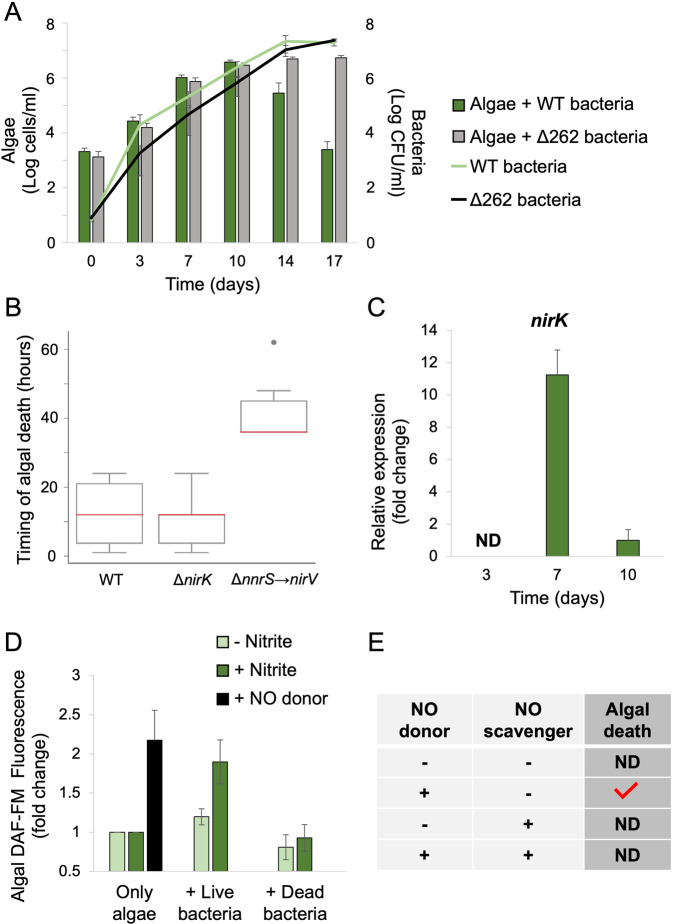
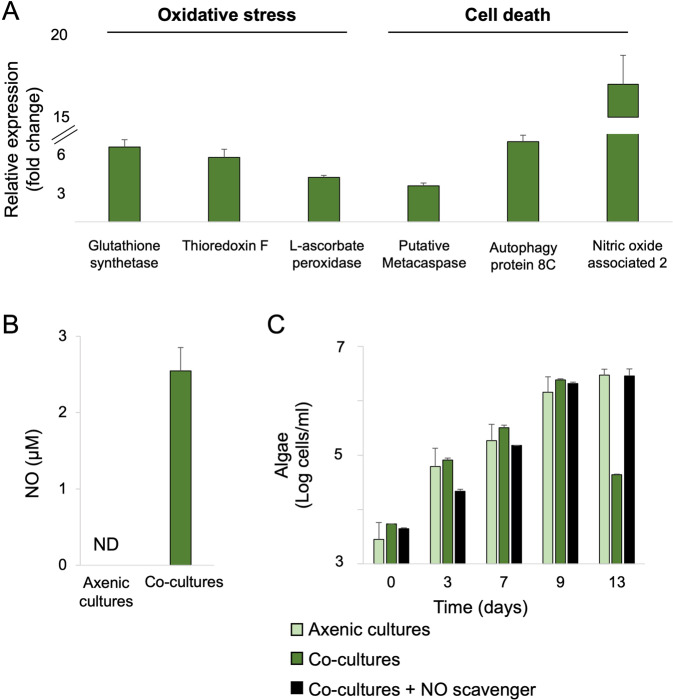
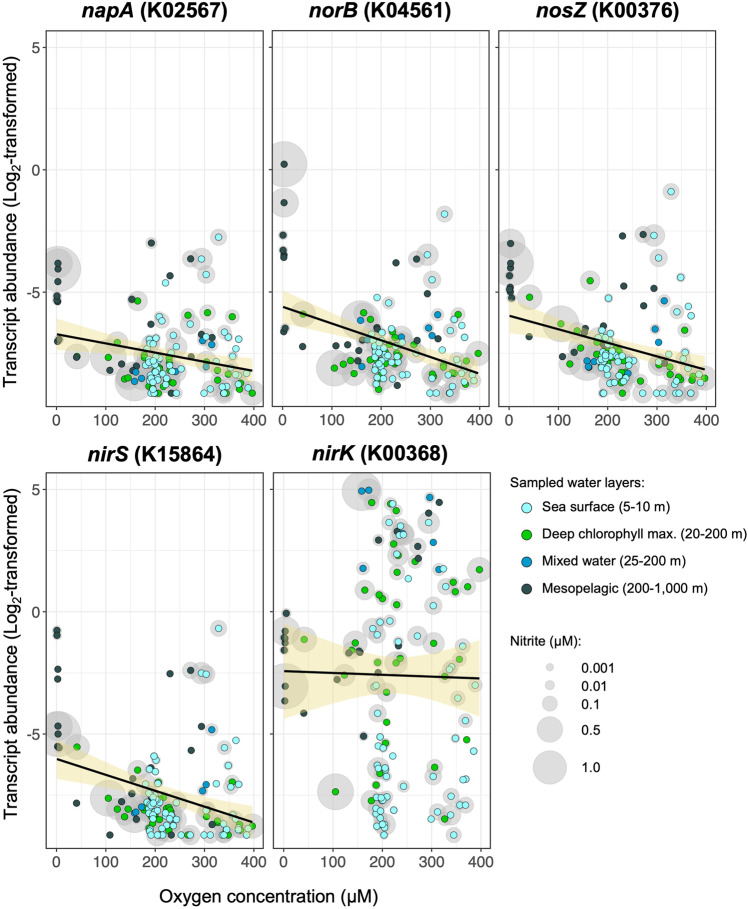
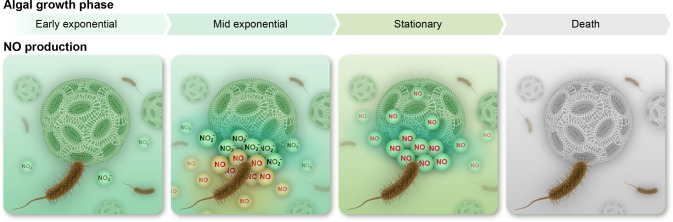
Microbial interactions govern marine biogeochemistry. These interactions are generally considered to rely on exchange of organic molecules. Here we report on a novel inorganic route of microbial communication, showing that algal-bacterial interactions between Phaeobacter inhibens bacteria and Gephyrocapsa huxleyi algae are mediated through inorganic nitrogen exchange. Under oxygen-rich conditions, aerobic bacteria reduce algal-secreted nitrite to nitric oxide (NO) through denitrification, a well-studied anaerobic respiratory mechanism. The bacterial NO is involved in triggering a cascade in algae akin to programmed cell death. During death, algae further generate NO, thereby propagating the signal in the algal population. Eventually, the algal population collapses, similar to the sudden demise of oceanic algal blooms. Our study suggests that the exchange of inorganic nitrogen species in oxygenated environments is a potentially significant route of microbial communication within and across kingdoms.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Jorgensen KS, Jensen HB, Sorensen J. Nitrous-oxide production from nitrification and denitrification in marine sediment at low oxygen concentrations. Can J Microbiol. 1984;30:1073–8.
-
- Ward BB, Zafiriou OC. Nitrification and nitric-oxide in the oxygen minimum of the eastern tropical north pacific. Deep-Sea Res. 1988;35:1127–42.
-
- Babbin AR, Bianchi D, Jayakumar A, Ward BB. Rapid nitrous oxide cycling in the suboxic ocean. Science. 2015;348:1127–9. - PubMed
-
- Babbin AR, Keil RG, Devol AH, Ward BB. Organic matter stoichiometry, flux, and oxygen control nitrogen loss in the ocean. Science. 2014;344:406–8. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
