

ROS Consumers or Producers? Interpreting Transcriptomic Data by AlphaFold Modeling Provides Insights into Class III Peroxidase Functions in Response to Biotic and Abiotic Stresses
- PMID: 37176003
- PMCID: PMC10179425
- DOI: 10.3390/ijms24098297
ROS Consumers or Producers? Interpreting Transcriptomic Data by AlphaFold Modeling Provides Insights into Class III Peroxidase Functions in Response to Biotic and Abiotic Stresses
Abstract
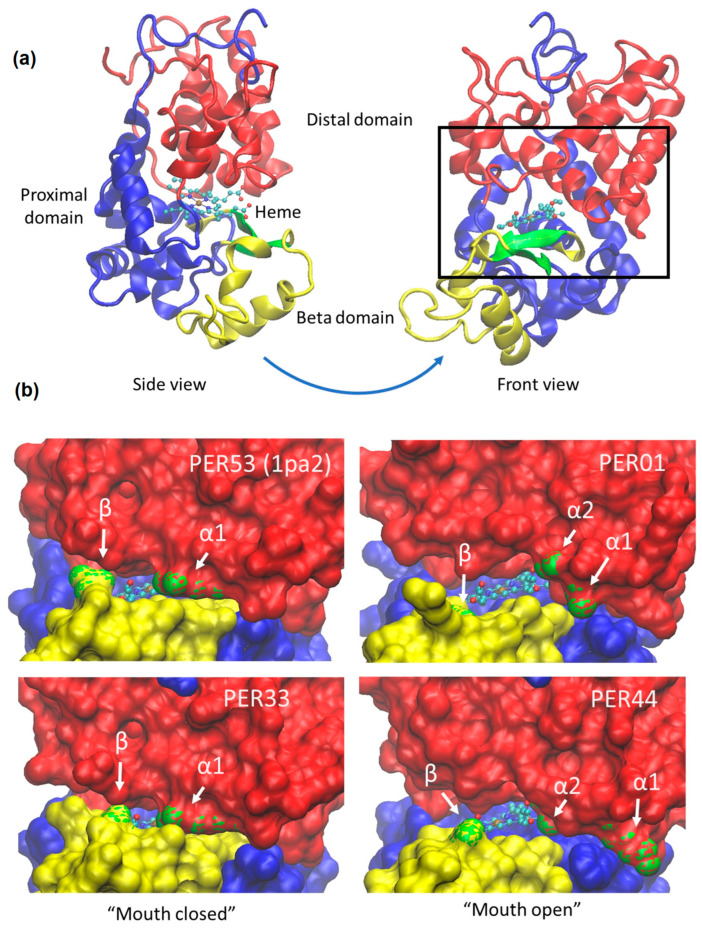
Participating in both biotic and abiotic stress responses, plant-specific class III peroxidases (PERs) show promise as candidates for crop improvement. The multigenic PER family is known to take part in diverse functions, such as lignin formation and defense against pathogens. Traditionally linked to hydrogen peroxide (H2O2) consumption, PERs can also produce reactive oxygen species (ROS), essential in tissue development, pathogen defense and stress signaling. The amino acid sequences of both orthologues and paralogues of PERs are highly conserved, but discovering correlations between sequence differences and their functional diversity has proven difficult. By combining meta-analysis of transcriptomic data and sequence alignments, we discovered a correlation between three key amino acid positions and gene expression in response to biotic and abiotic stresses. Phylogenetic analysis revealed evolutionary pressure on these amino acids toward stress responsiveness. Using AlphaFold modeling, we found unique interdomain and protein-heme interactions involving those key amino acids in stress-induced PERs. Plausibly, these structural interactions may act as "gate keepers" by preventing larger substrates from accessing the heme and thereby shifting PER function from consumption to the production of ROS.
Keywords: AlphaFold; Arabidopsis; ROS signaling; class III peroxidases; phosphate deficiency; plant stress response.
Conflict of interest statement
The authors declare no conflict of interest.
Figures



References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
