

FRA-1 as a Regulator of EMT and Metastasis in Breast Cancer
- PMID: 37176013
- PMCID: PMC10179602
- DOI: 10.3390/ijms24098307
FRA-1 as a Regulator of EMT and Metastasis in Breast Cancer
Abstract
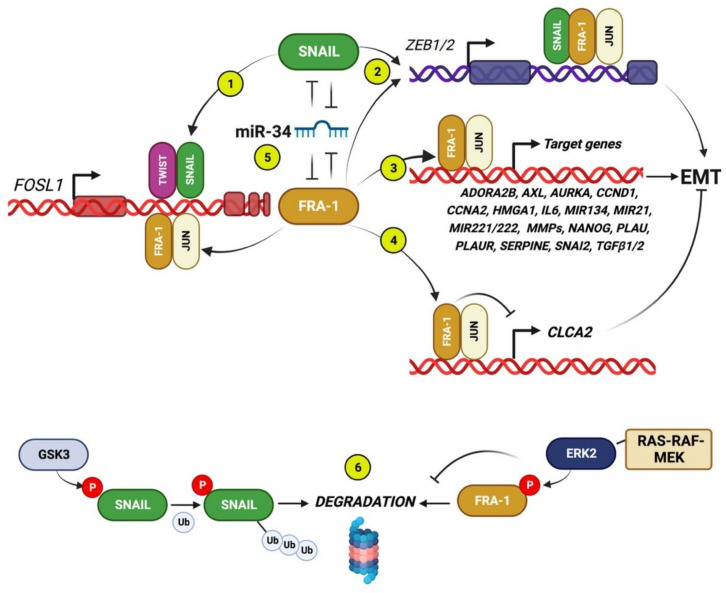
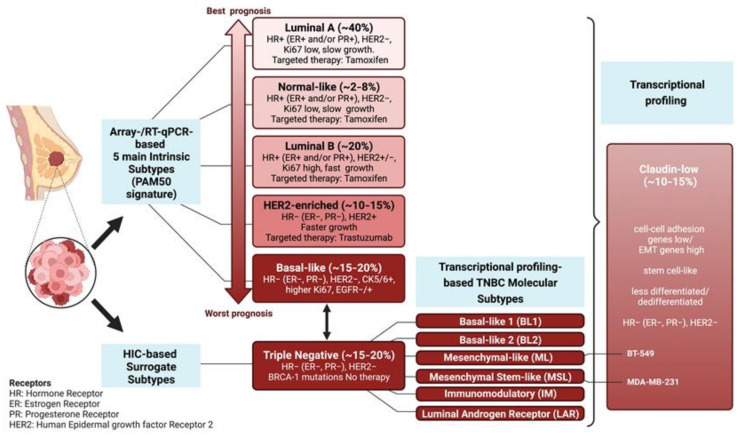
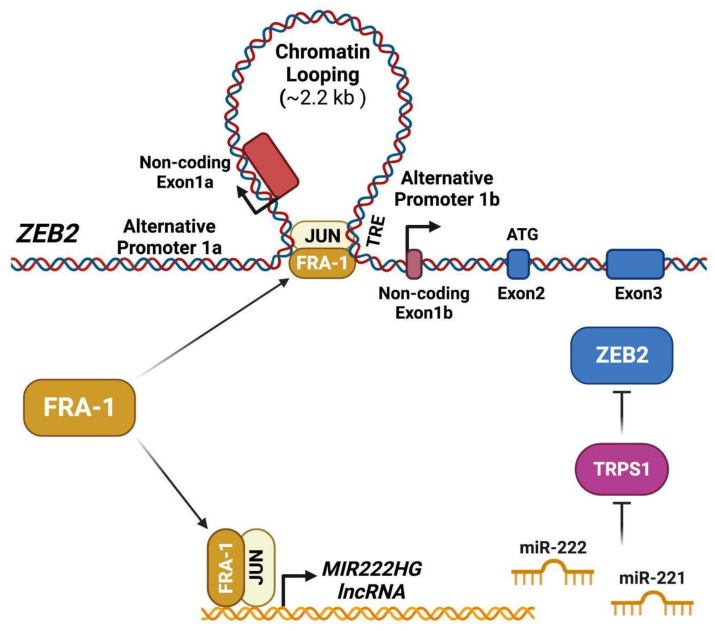
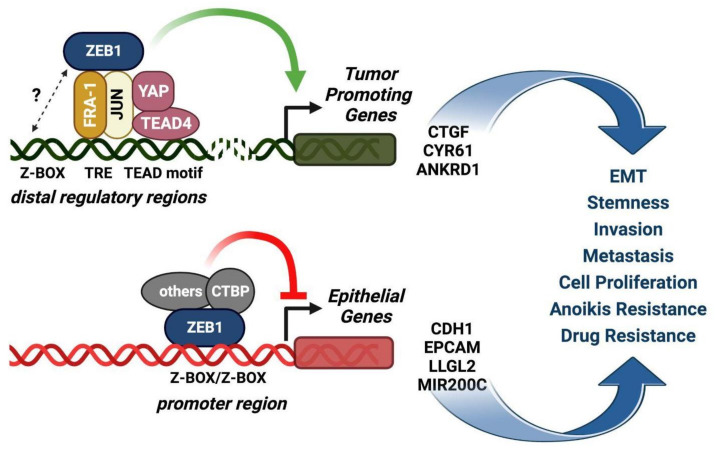
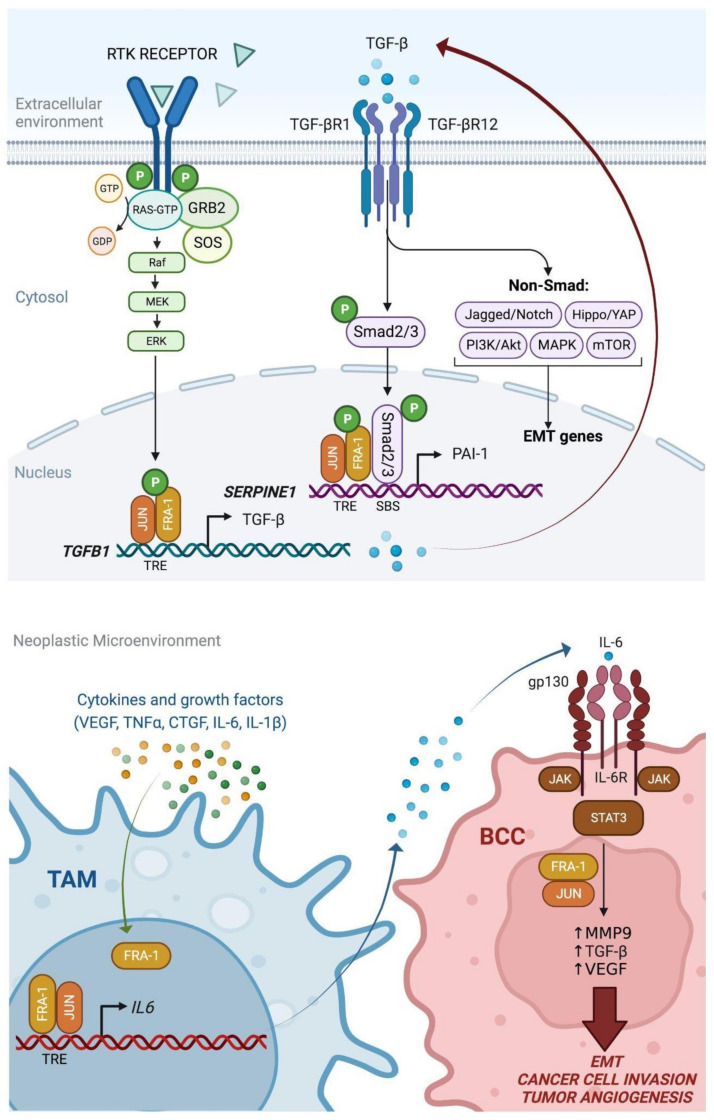
Among FOS-related components of the dimeric AP-1 transcription factor, the oncoprotein FRA-1 (encoded by FOSL1) is a key regulator of invasion and metastasis. The well-established FRA-1 pro-invasive activity in breast cancer, in which FOSL1 is overexpressed in the TNBC (Triple Negative Breast Cancer)/basal subtypes, correlates with the FRA-1-dependent transcriptional regulation of EMT (Epithelial-to-Mesenchymal Transition). After summarizing the major findings on FRA-1 in breast cancer invasiveness, we discuss the FRA-1 mechanistic links with EMT and cancer cell stemness, mediated by transcriptional and posttranscriptional interactions between FOSL1/FRA-1 and EMT-regulating transcription factors, miRNAs, RNA binding proteins and cytokines, along with other target genes involved in EMT. In addition to the FRA-1/AP-1 effects on the architecture of target promoters, we discuss the diagnostic and prognostic significance of the EMT-related FRA-1 transcriptome, along with therapeutic implications. Finally, we consider several novel perspectives regarding the less explored roles of FRA-1 in the tumor microenvironment and in control of the recently characterized hybrid EMT correlated with cancer cell plasticity, stemness, and metastatic potential. We will also examine the application of emerging technologies, such as single-cell analyses, along with animal models of TNBC and tumor-derived CTCs and PDXs (Circulating Tumor Cells and Patient-Derived Xenografts) for studying the FRA-1-mediated mechanisms in in vivo systems of EMT and metastasis.
Keywords: AP-1 transcription factors; EMT; FOSL1; FRA-1; TNBC.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
The nuclear oncoprotein Fra-1: a transcription factor knocking on therapeutic applications' door.Oncogene. 2020 Jun;39(23):4491-4506. doi: 10.1038/s41388-020-1306-4. Epub 2020 May 8. Oncogene. 2020. PMID: 32385348 Review.
-
Fra-1/AP-1 induces EMT in mammary epithelial cells by modulating Zeb1/2 and TGFβ expression.Cell Death Differ. 2015 Feb;22(2):336-50. doi: 10.1038/cdd.2014.157. Epub 2014 Oct 10. Cell Death Differ. 2015. PMID: 25301070 Free PMC article.
-
The Fra-1/AP-1 Oncoprotein: From the "Undruggable" Transcription Factor to Therapeutic Targeting.Cancers (Basel). 2022 Mar 14;14(6):1480. doi: 10.3390/cancers14061480. Cancers (Basel). 2022. PMID: 35326630 Free PMC article. Review.
-
Aberrantly expressed Fra-1 by IL-6/STAT3 transactivation promotes colorectal cancer aggressiveness through epithelial-mesenchymal transition.Carcinogenesis. 2015 Apr;36(4):459-68. doi: 10.1093/carcin/bgv017. Epub 2015 Mar 6. Carcinogenesis. 2015. PMID: 25750173 Free PMC article.
-
miR-124 regulates EMT based on ZEB2 target to inhibit invasion and metastasis in triple-negative breast cancer.Pathol Res Pract. 2019 Apr;215(4):697-704. doi: 10.1016/j.prp.2018.12.039. Epub 2018 Dec 31. Pathol Res Pract. 2019. PMID: 30611621
Cited by
-
Fra-1 affects chemotherapy sensitivity by inhibiting ferroptosis in gastric cancer cells.Cancer Drug Resist. 2024 Nov 16;7:44. doi: 10.20517/cdr.2024.101. eCollection 2024. Cancer Drug Resist. 2024. PMID: 39624082 Free PMC article.
-
CRISPR/Cas9 as a therapeutic tool for triple negative breast cancer: from bench to clinics.Front Mol Biosci. 2023 Jul 4;10:1214489. doi: 10.3389/fmolb.2023.1214489. eCollection 2023. Front Mol Biosci. 2023. PMID: 37469704 Free PMC article. Review.
-
FOSL1 transcriptionally dictates the Warburg effect and enhances chemoresistance in triple-negative breast cancer.J Transl Med. 2025 Jan 2;23(1):1. doi: 10.1186/s12967-024-06014-9. J Transl Med. 2025. PMID: 39748430 Free PMC article.
-
Noncoding RNAs as regulators of FOSL1 in cancer.Front Immunol. 2025 Aug 1;16:1599674. doi: 10.3389/fimmu.2025.1599674. eCollection 2025. Front Immunol. 2025. PMID: 40821785 Free PMC article. Review.
-
FOSL1 is a key regulator of a super-enhancer driving TCOF1 expression in triple-negative breast cancer.Epigenetics Chromatin. 2024 Nov 10;17(1):34. doi: 10.1186/s13072-024-00559-1. Epigenetics Chromatin. 2024. PMID: 39523372 Free PMC article.
References
-
- Sørlie T., Perou C.M., Tibshirani R., Aas T., Geisler S., Johnsen H., Hastie T., Eisen M.B., Van De Rijn M., Jeffrey S.S., et al. Gene expression patterns of breast carcinomas distinguish tumor subclasses with clinical implications. Proc. Natl. Acad. Sci. USA. 2001;98:10869–10874. doi: 10.1073/pnas.191367098. - DOI - PMC - PubMed
-
- DeVita V.T., Lawrence T.S., Rosenberg S.A. Cancer: Principles & Practice of Oncology: Primer of the Molecular Biology of Cancer. Wolters Kluwer Health/Lippincott Williams & Wilkins; Philadelphia, PA, USA: 2011.
-
- Van Dam P.-J., Van Laere S. Molecular profiling in cancer research and personalized medicine. In: Pezzella F., Tavassoli M., Kerr D.J., editors. Oxford Textbook of Cancer Biology. Oxford University Press; Oxford, UK: 2019. pp. 347–362. - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
