

DNA modifications impact natural transformation of Acinetobacter baumannii
- PMID: 37178001
- PMCID: PMC10287943
- DOI: 10.1093/nar/gkad377
DNA modifications impact natural transformation of Acinetobacter baumannii
Abstract
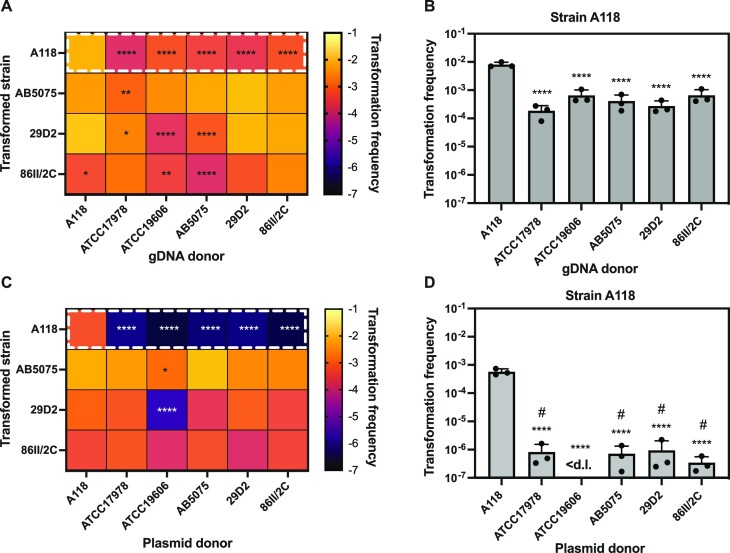
Acinetobacter baumannii is a dangerous nosocomial pathogen, especially due to its ability to rapidly acquire new genetic traits, including antibiotic resistance genes (ARG). In A. baumannii, natural competence for transformation, one of the primary modes of horizontal gene transfer (HGT), is thought to contribute to ARG acquisition and has therefore been intensively studied. However, knowledge regarding the potential role of epigenetic DNA modification(s) on this process remains lacking. Here, we demonstrate that the methylome pattern of diverse A. baumannii strains differs substantially and that these epigenetic marks influence the fate of transforming DNA. Specifically, we describe a methylome-dependent phenomenon that impacts intra- and inter-species DNA exchange by the competent A. baumannii strain A118. We go on to identify and characterize an A118-specific restriction-modification (RM) system that impairs transformation when the incoming DNA lacks a specific methylation signature. Collectively, our work contributes towards a more holistic understanding of HGT in this organism and may also aid future endeavors towards tackling the spread of novel ARGs. In particular, our results suggest that DNA exchanges between bacteria that share similar epigenomes are favored and could therefore guide future research into identifying the reservoir(s) of dangerous genetic traits for this multi-drug resistant pathogen.
© The Author(s) 2023. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures





Comment in
-
Restriction-methylation systems regulate transformation in Acinetobacter baumannii.Trends Microbiol. 2023 Sep;31(9):879-881. doi: 10.1016/j.tim.2023.07.009. Epub 2023 Jul 24. Trends Microbiol. 2023. PMID: 37495486
Similar articles
-
Pilus Production in Acinetobacter baumannii Is Growth Phase Dependent and Essential for Natural Transformation.J Bacteriol. 2021 Mar 23;203(8):e00034-21. doi: 10.1128/JB.00034-21. Print 2021 Mar 23. J Bacteriol. 2021. PMID: 33495250 Free PMC article.
-
Serum Albumin and Ca2+ Are Natural Competence Inducers in the Human Pathogen Acinetobacter baumannii.Antimicrob Agents Chemother. 2016 Jul 22;60(8):4920-9. doi: 10.1128/AAC.00529-16. Print 2016 Aug. Antimicrob Agents Chemother. 2016. PMID: 27270286 Free PMC article.
-
Restriction-methylation systems regulate transformation in Acinetobacter baumannii.Trends Microbiol. 2023 Sep;31(9):879-881. doi: 10.1016/j.tim.2023.07.009. Epub 2023 Jul 24. Trends Microbiol. 2023. PMID: 37495486
-
Carbapenemases: Transforming Acinetobacter baumannii into a Yet More Dangerous Menace.Biomolecules. 2020 May 6;10(5):720. doi: 10.3390/biom10050720. Biomolecules. 2020. PMID: 32384624 Free PMC article. Review.
-
Acinetobacter baumannii: evolution of antimicrobial resistance-treatment options.Semin Respir Crit Care Med. 2015 Feb;36(1):85-98. doi: 10.1055/s-0034-1398388. Epub 2015 Feb 2. Semin Respir Crit Care Med. 2015. PMID: 25643273 Free PMC article. Review.
Cited by
-
Generation of tools for expression and purification of the phage-encoded Type I restriction enzyme inhibitor, Ocr.Microbiology (Reading). 2024 Jul;170(7):001465. doi: 10.1099/mic.0.001465. Microbiology (Reading). 2024. PMID: 39042422 Free PMC article.
-
Advances in Diversity, Evolutionary Dynamics and Biotechnological Potential of Restriction-Modification Systems.Microorganisms. 2025 May 14;13(5):1126. doi: 10.3390/microorganisms13051126. Microorganisms. 2025. PMID: 40431298 Free PMC article. Review.
-
Intragenomic conflicts with plasmids and chromosomal mobile genetic elements drive the evolution of natural transformation within species.PLoS Biol. 2024 Oct 14;22(10):e3002814. doi: 10.1371/journal.pbio.3002814. eCollection 2024 Oct. PLoS Biol. 2024. PMID: 39401218 Free PMC article.
-
Large DNA fragment ISEc9-mediated transposition during natural transformation allows interspecies dissemination of antimicrobial resistance genes.Eur J Clin Microbiol Infect Dis. 2025 Jun;44(6):1417-1424. doi: 10.1007/s10096-025-05113-9. Epub 2025 Mar 28. Eur J Clin Microbiol Infect Dis. 2025. PMID: 40304893 Free PMC article.
-
DNA binding is rate-limiting for natural transformation.bioRxiv [Preprint]. 2024 Jun 6:2024.06.06.597730. doi: 10.1101/2024.06.06.597730. bioRxiv. 2024. PMID: 38895488 Free PMC article. Preprint.
References
-
- Rice L.B. Federal funding for the study of antimicrobial resistance in nosocomial pathogens: no ESKAPE. J. Infect. Dis. 2008; 197:1079–1081. - PubMed
-
- Shrestha S., Tada T., Miyoshi-Akiyama T., Ohara H., Shimada K., Satou K., Teruya K., Nakano K., Shiroma A., Sherchand J.B.et al. .. Molecular epidemiology of multidrug-resistant Acinetobacterbaumannii isolates in a university hospital in Nepal reveals the emergence of a novel epidemic clonal lineage. Int. J. Antimicrob. Agents. 2015; 46:526–531. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
