

Reliable and robust control of nucleus centering is contingent on nonequilibrium force patterns
- PMID: 37182105
- PMCID: PMC10173738
- DOI: 10.1016/j.isci.2023.106665
Reliable and robust control of nucleus centering is contingent on nonequilibrium force patterns
Abstract
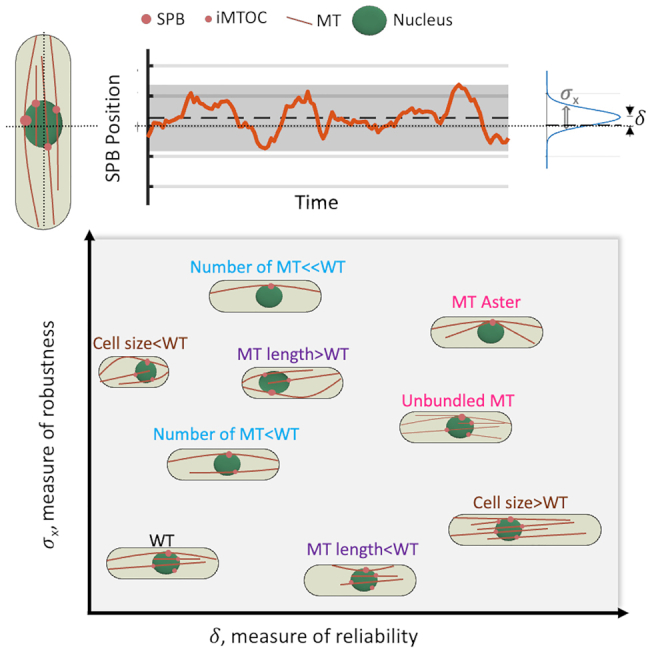
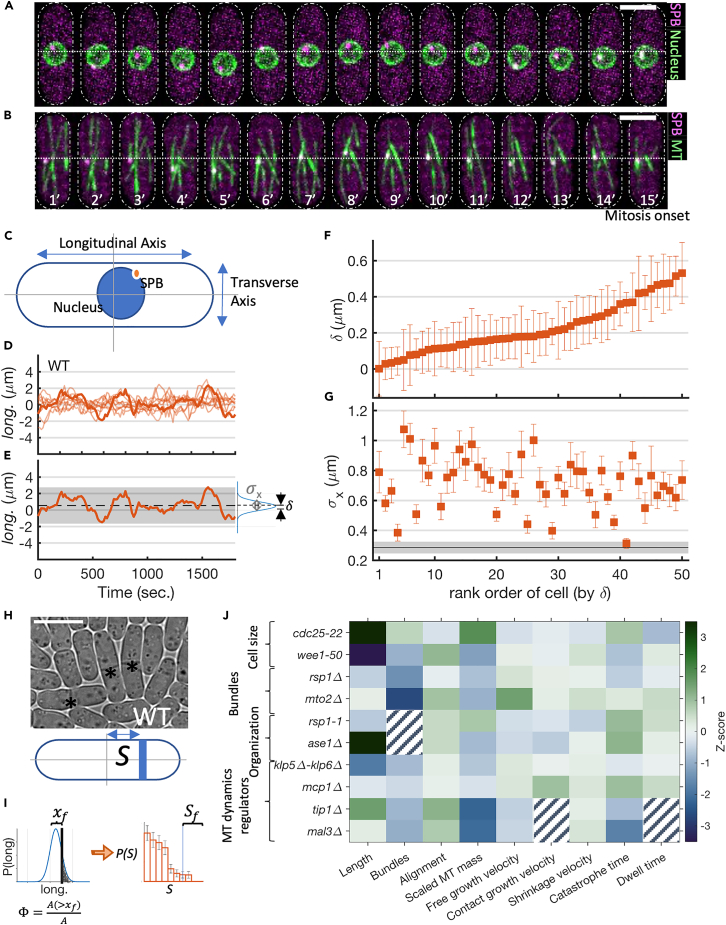
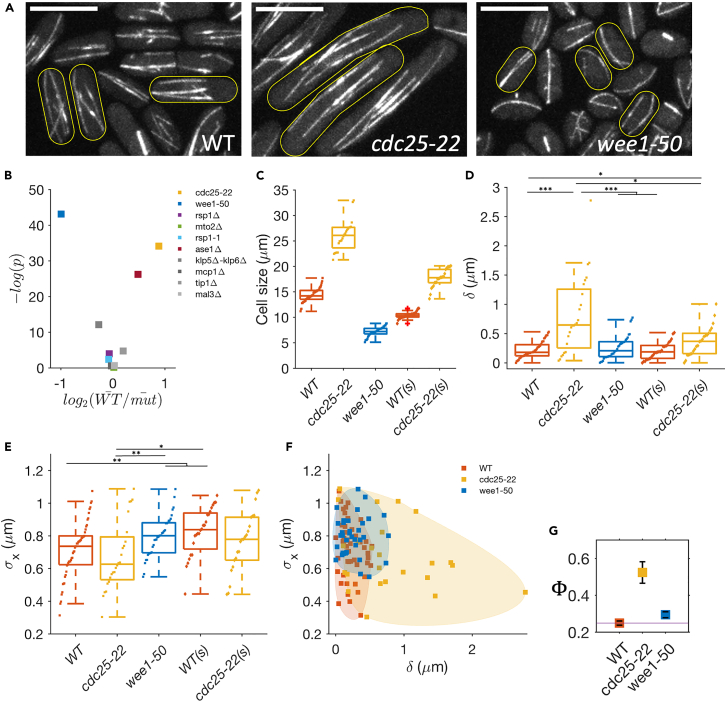
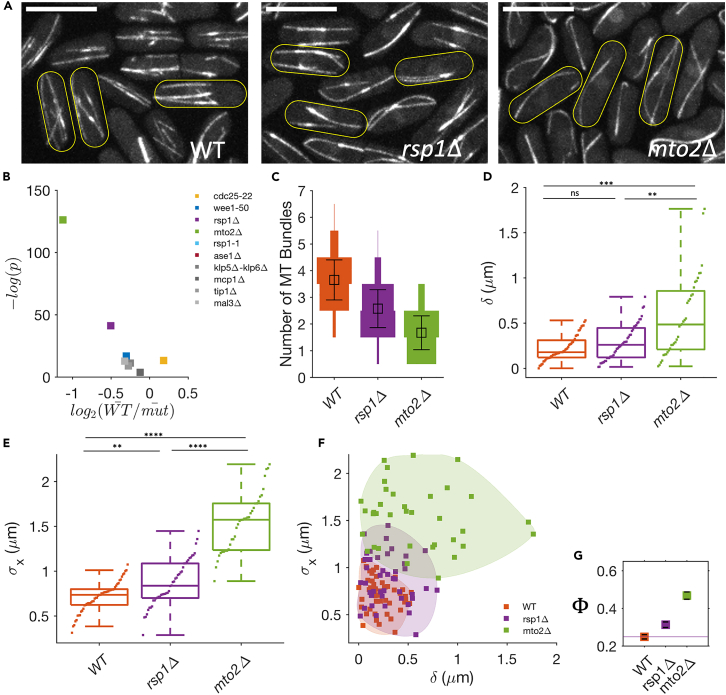
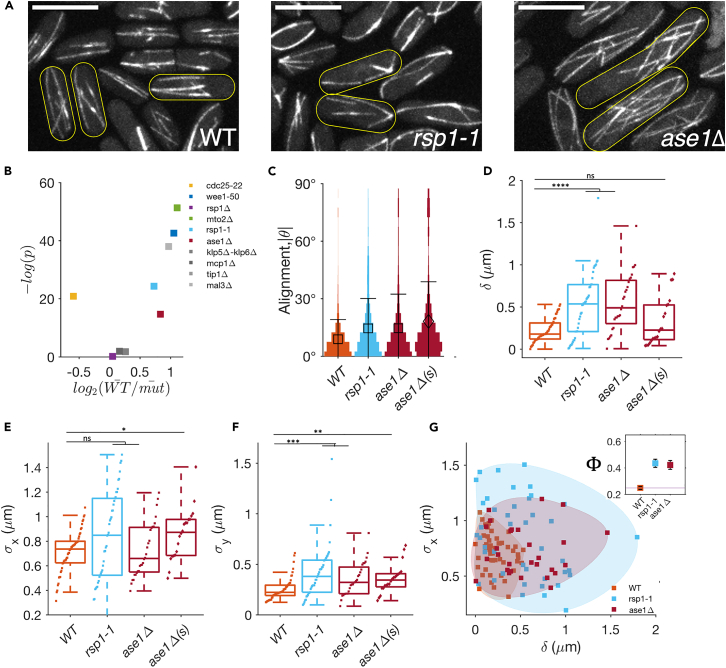
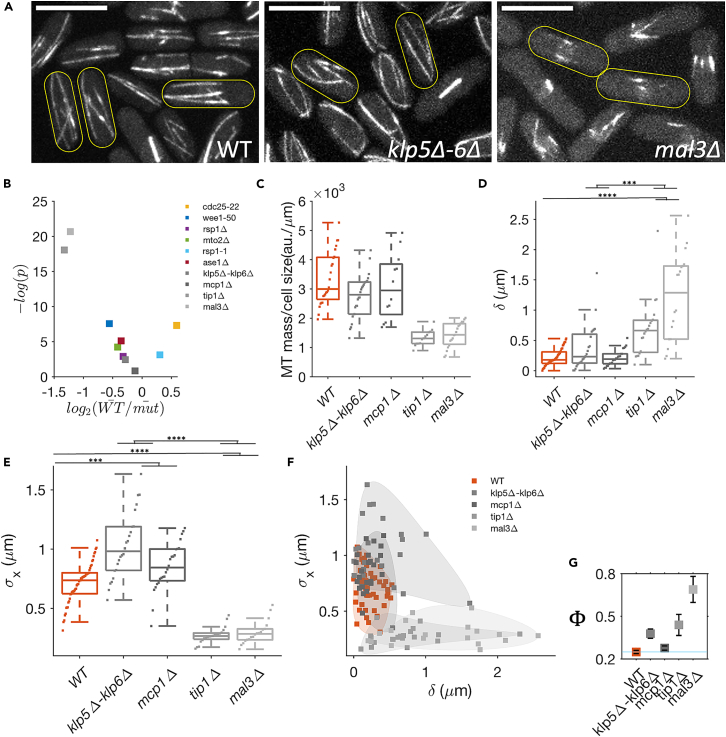
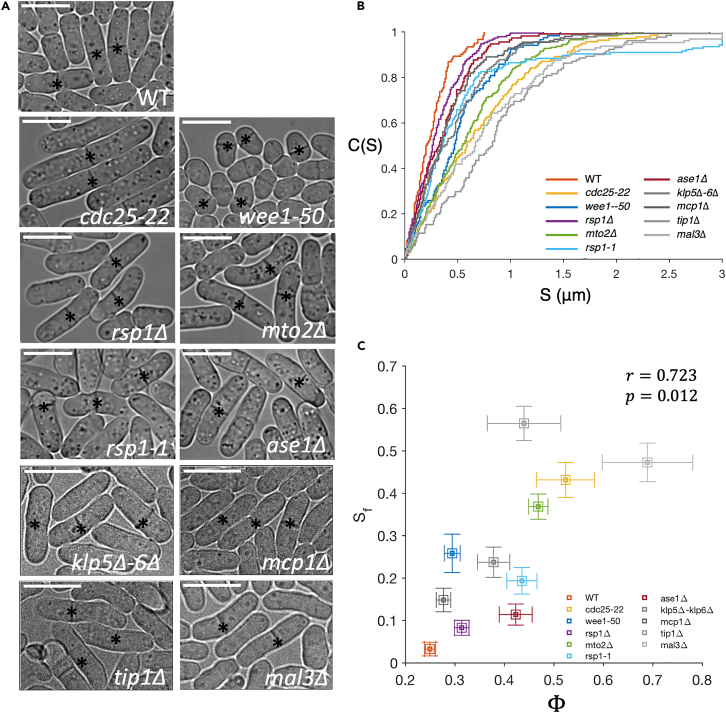
Cell centers their division apparatus to ensure symmetric cell division, a challenging task when the governing dynamics is stochastic. Using fission yeast, we show that the patterning of nonequilibrium polymerization forces of microtubule (MT) bundles controls the precise localization of spindle pole body (SPB), and hence the division septum, at the onset of mitosis. We define two cellular objectives, reliability, the mean SPB position relative to the geometric center, and robustness, the variance of the SPB position, which are sensitive to genetic perturbations that change cell length, MT bundle number/orientation, and MT dynamics. We show that simultaneous control of reliability and robustness is required to minimize septum positioning error achieved by the wild type (WT). A stochastic model for the MT-based nucleus centering, with parameters measured directly or estimated using Bayesian inference, recapitulates the maximum fidelity of WT. Using this, we perform a sensitivity analysis of the parameters that control nuclear centering.
Keywords: Biological sciences; Microbiology; Molecular biology; Molecular microbiology.
© 2023 The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures

Similar articles
-
Collective dynein transport of the nucleus by pulling on astral microtubules during Saccharomyces cerevisiae mitosis.Yeast. 2021 Jun;38(6):352-366. doi: 10.1002/yea.3552. Epub 2021 Feb 27. Yeast. 2021. PMID: 33547820
-
Microtubule dynamics from mating through the first zygotic division in the budding yeast Saccharomyces cerevisiae.J Cell Biol. 1999 Mar 8;144(5):977-87. doi: 10.1083/jcb.144.5.977. J Cell Biol. 1999. PMID: 10085295 Free PMC article.
-
Asymmetric microtubule pushing forces in nuclear centering.Curr Biol. 2006 Aug 8;16(15):1544-50. doi: 10.1016/j.cub.2006.06.026. Curr Biol. 2006. PMID: 16890530
-
Regulation of centrosome separation in yeast and vertebrates: common threads.Trends Cell Biol. 2009 Jul;19(7):325-33. doi: 10.1016/j.tcb.2009.03.008. Epub 2009 Jul 1. Trends Cell Biol. 2009. PMID: 19576775 Review.
-
Duplication and Nuclear Envelope Insertion of the Yeast Microtubule Organizing Centre, the Spindle Pole Body.Cells. 2018 May 10;7(5):42. doi: 10.3390/cells7050042. Cells. 2018. PMID: 29748517 Free PMC article. Review.
Cited by
-
Force-transducing molecular ensembles at growing microtubule tips control mitotic spindle size.Nat Commun. 2024 Nov 14;15(1):9865. doi: 10.1038/s41467-024-54123-2. Nat Commun. 2024. PMID: 39543105 Free PMC article.
References
LinkOut - more resources
Full Text Sources
