

Cell entry and release of quasi-enveloped human hepatitis viruses
- PMID: 37185947
- PMCID: PMC10127183
- DOI: 10.1038/s41579-023-00889-z
Cell entry and release of quasi-enveloped human hepatitis viruses
Abstract
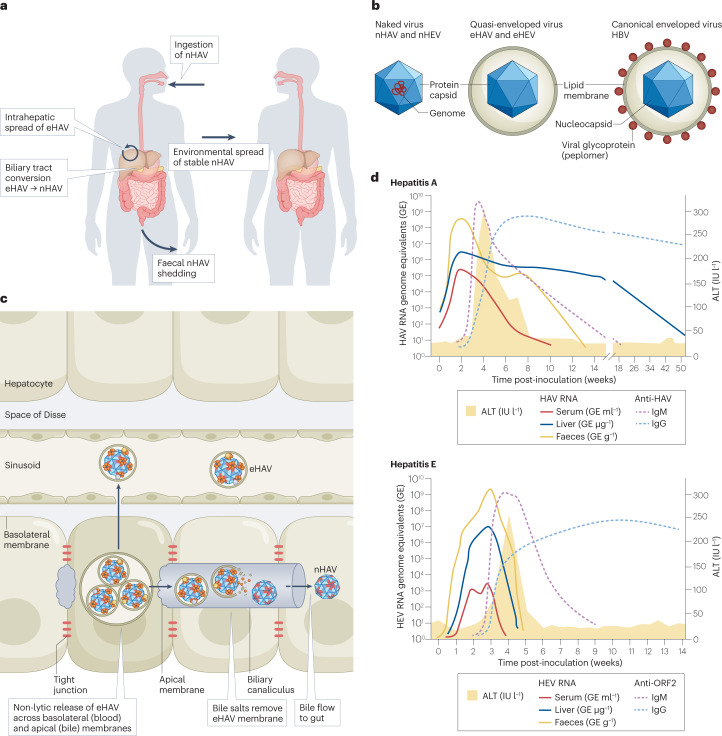
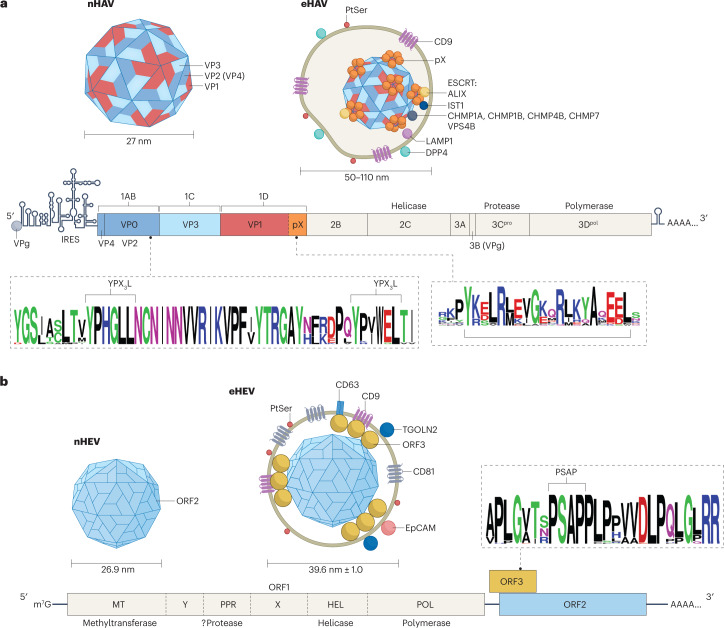
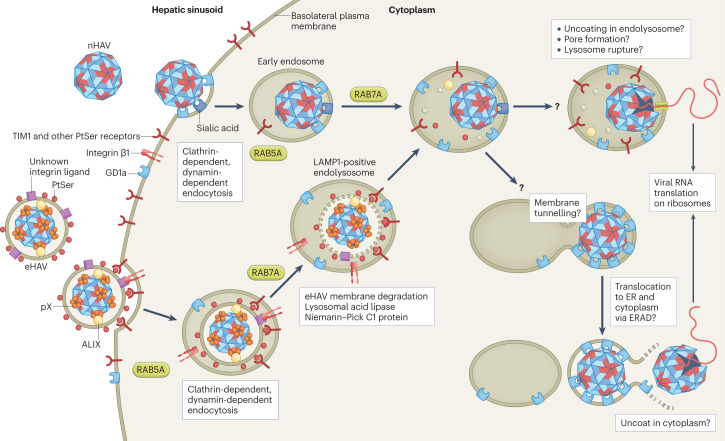
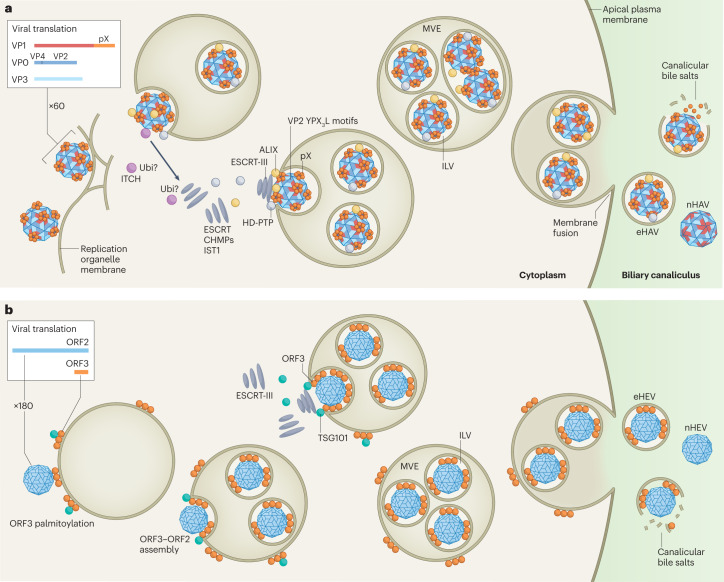
Infectious hepatitis type A and type E are caused by phylogenetically distinct single-stranded, positive-sense RNA viruses that were once considered to be non-enveloped. However, studies show that both are released nonlytically from hepatocytes as 'quasi-enveloped' virions cloaked in host membranes. These virion types predominate in the blood of infected individuals and mediate virus spread within the liver. They lack virally encoded proteins on their surface and are resistant to neutralizing anti-capsid antibodies induced by infection, yet they efficiently enter cells and initiate new rounds of virus replication. In this Review, we discuss the mechanisms by which specific peptide sequences in the capsids of these quasi-enveloped virions mediate their endosomal sorting complexes required for transport (ESCRT)-dependent release from hepatocytes through multivesicular endosomes, what is known about how they enter cells, and the impact of capsid quasi-envelopment on host immunity and pathogenesis.
© 2023. Springer Nature Limited.
Conflict of interest statement
SML is co-inventor on a pending patent application related to antiviral compounds with activity against hepatitis A virus. The other authors have no competing interests to declare.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
