

FGF21 has a sex-specific role in calorie-restriction-induced beiging of white adipose tissue in mice
- PMID: 37186544
- PMCID: PMC10181818
FGF21 has a sex-specific role in calorie-restriction-induced beiging of white adipose tissue in mice
Abstract
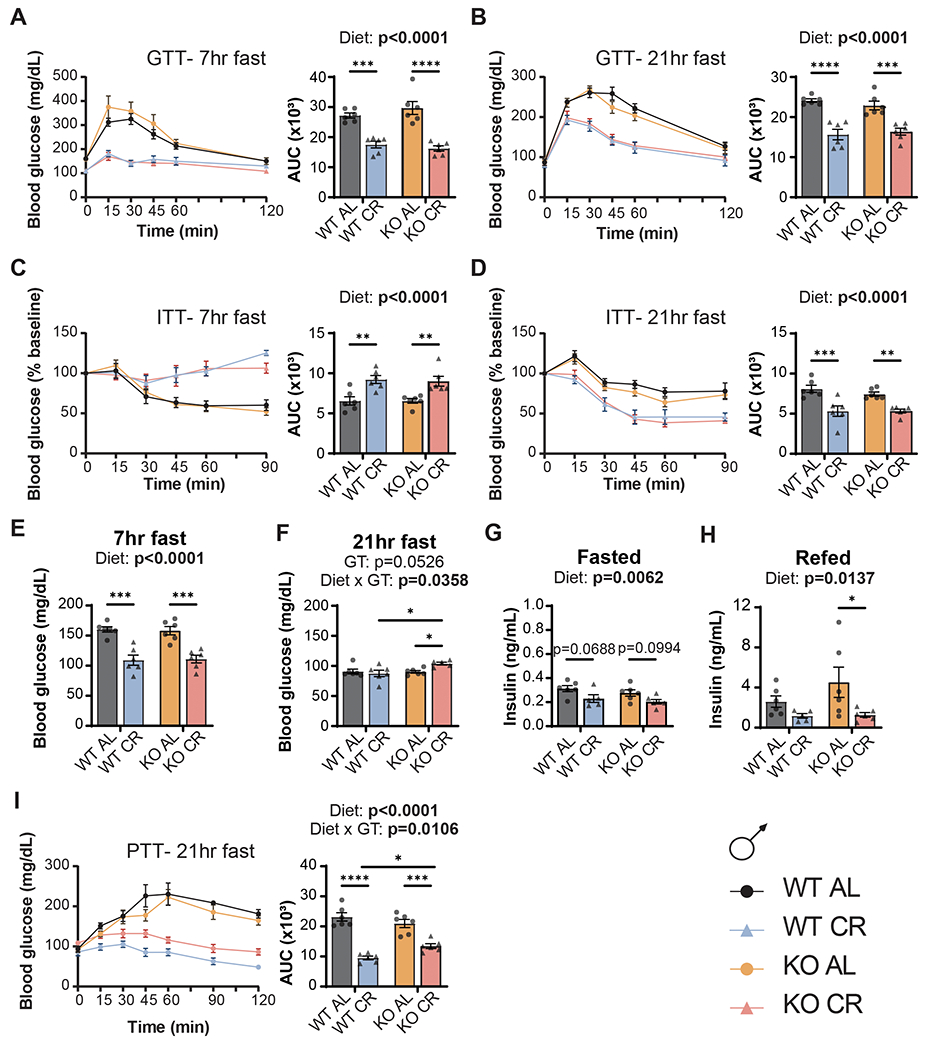
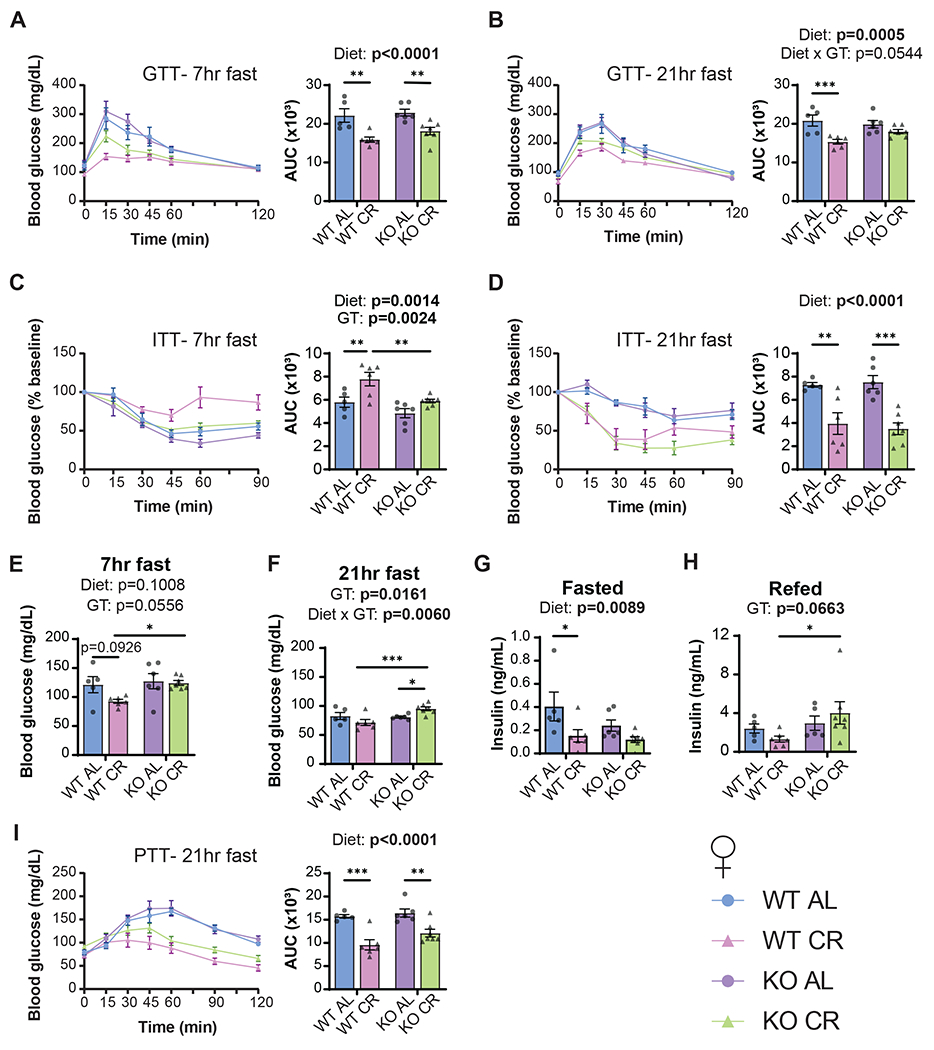
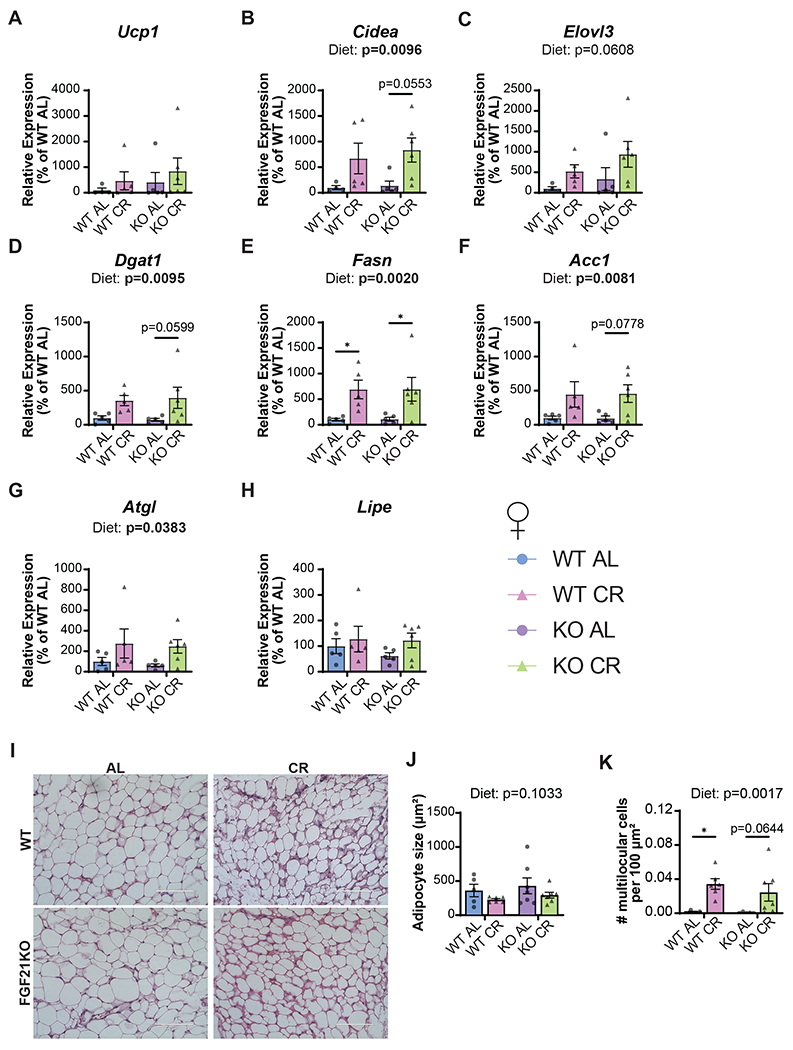
Calorie restriction (CR) promotes healthspan and extends the lifespan of diverse organisms, including mice, and there is intense interest in understanding the molecular mechanisms by which CR functions. Some studies have demonstrated that CR induces fibroblast growth factor 21 (FGF21), a hormone that regulates energy balance and that when overexpressed, promotes metabolic health and longevity in mice, but the role of FGF21 in the response to CR has not been fully investigated. We directly examined the role of FGF21 in the physiological and metabolic response to a CR diet by feeding Fgf21-/- and wild-type control mice either ad libitum (AL) diet or a 30% CR diet for 15 weeks. Here, we find that FGF21 is largely dispensable for CR-induced improvements in body composition and energy balance, but that lack of Fgf21 blunts CR-induced changes aspects of glucose regulation and insulin sensitivity in females. Surprisingly, despite not affecting CR-induced changes in energy expenditure, loss of Fgf21 significantly blunts CR-induced beiging of white adipose tissue in male but not female mice. Our results shed new light on the molecular mechanisms involved in the beneficial effects of a CR diet, clarify that FGF21 is largely dispensable for the metabolic effects of a CR diet, and highlight a sex-dependent role for FGF21 in the molecular adaptation of white adipose tissue to CR.
Keywords: FGF21; beiging; calorie restriction; glucose homeostasis; metabolic health; white adipose tissue.
Conflict of interest statement
D.W.L has received funding from, and is a scientific advisory board member of, Aeovian Pharmaceuticals, which seeks to develop novel, selective mTOR inhibitors for the treatment of various diseases. The remaining authors declare no competing interests.
Figures





References
Grants and funding
LinkOut - more resources
Full Text Sources