

α-Synuclein fibril and synaptic vesicle interactions lead to vesicle destruction and increased lipid-associated fibril uptake into iPSC-derived neurons
- PMID: 37188797
- PMCID: PMC10185682
- DOI: 10.1038/s42003-023-04884-1
α-Synuclein fibril and synaptic vesicle interactions lead to vesicle destruction and increased lipid-associated fibril uptake into iPSC-derived neurons
Abstract
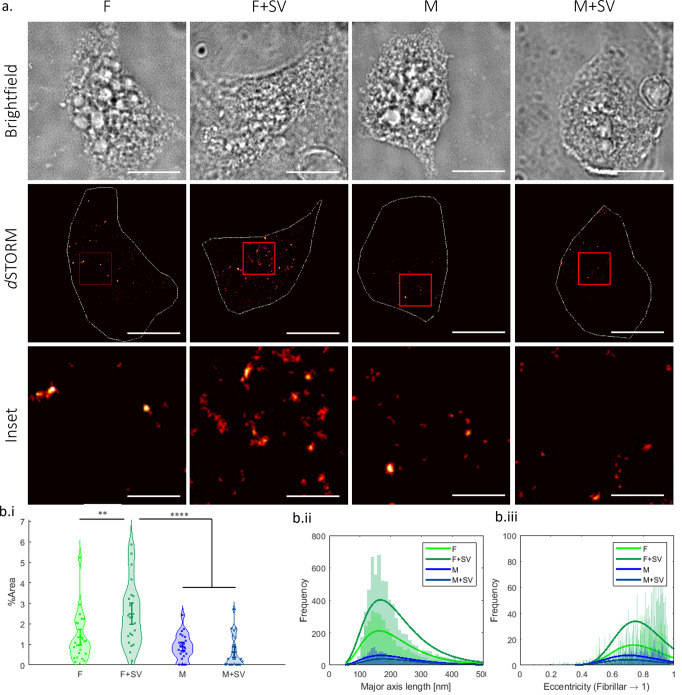
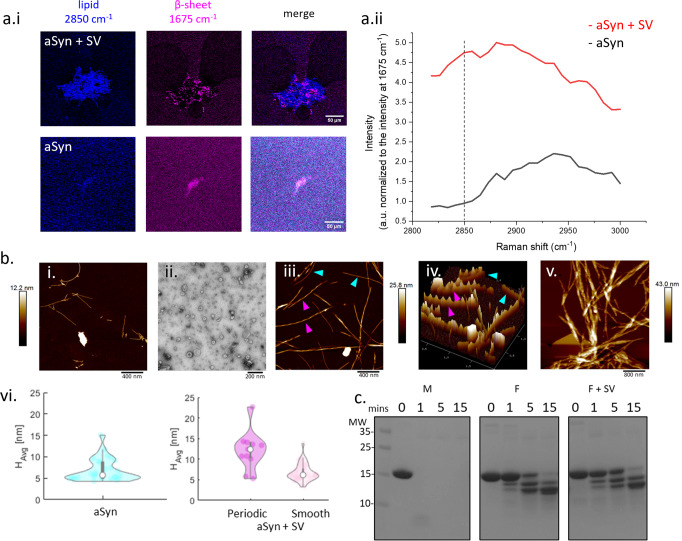
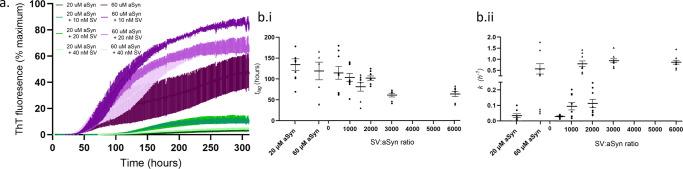
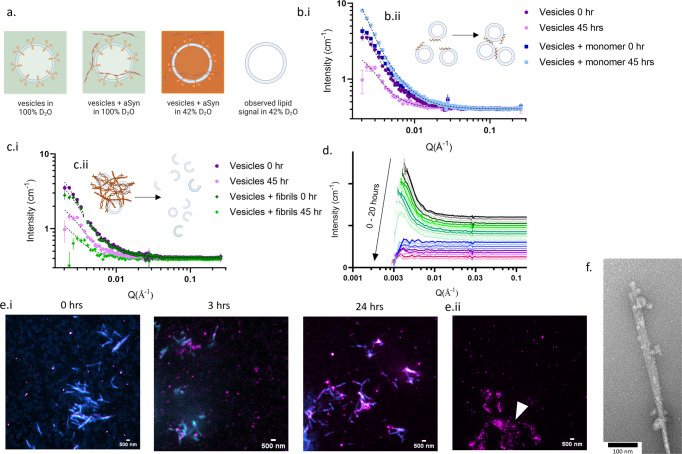
Monomeric alpha-synuclein (aSyn) is a well characterised protein that importantly binds to lipids. aSyn monomers assemble into amyloid fibrils which are localised to lipids and organelles in insoluble structures found in Parkinson's disease patient's brains. Previous work to address pathological aSyn-lipid interactions has focused on using synthetic lipid membranes, which lack the complexity of physiological lipid membranes. Here, we use physiological membranes in the form of synaptic vesicles (SV) isolated from rodent brain to demonstrate that lipid-associated aSyn fibrils are more easily taken up into iPSC-derived cortical i3Neurons. Lipid-associated aSyn fibril characterisation reveals that SV lipids are an integrated part of the fibrils and while their fibril morphology differs from aSyn fibrils alone, the core fibril structure remains the same, suggesting the lipids lead to the increase in fibril uptake. Furthermore, SV enhance the aggregation rate of aSyn, yet increasing the SV:aSyn ratio causes a reduction in aggregation propensity. We finally show that aSyn fibrils disintegrate SV, whereas aSyn monomers cause clustering of SV using small angle neutron scattering and high-resolution imaging. Disease burden on neurons may be impacted by an increased uptake of lipid-associated aSyn which could enhance stress and pathology, which in turn may have fatal consequences for neurons.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
