

Insights on Host-Parasite Immunomodulation Mediated by Extracellular Vesicles of Cutaneous Leishmania shawi and Leishmania guyanensis
- PMID: 37190011
- PMCID: PMC10137031
- DOI: 10.3390/cells12081101
Insights on Host-Parasite Immunomodulation Mediated by Extracellular Vesicles of Cutaneous Leishmania shawi and Leishmania guyanensis
Abstract
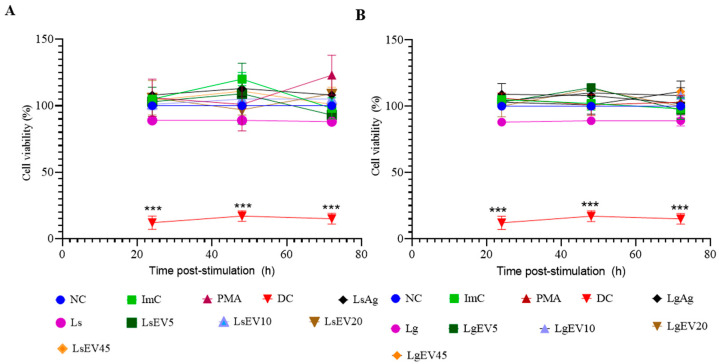
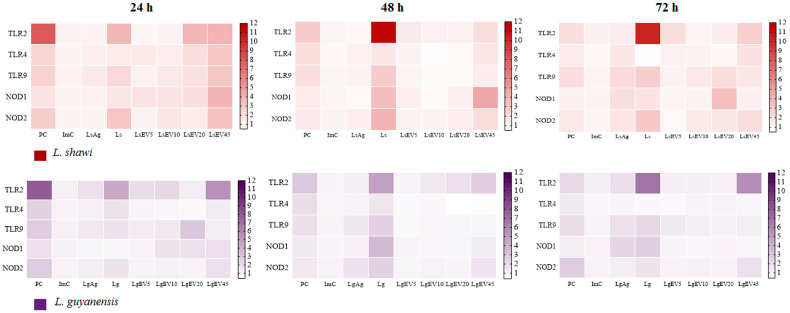
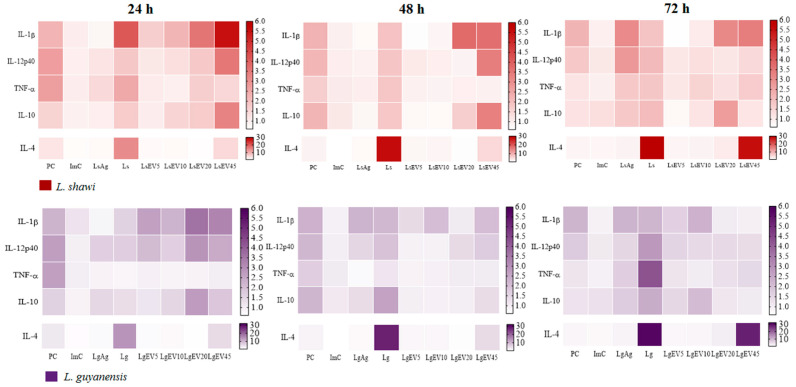
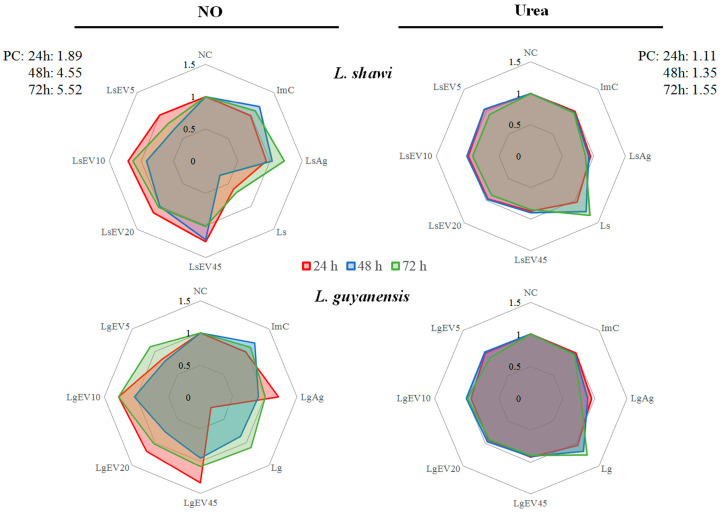
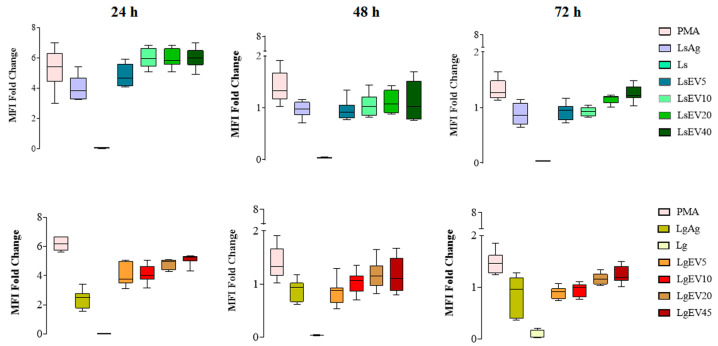
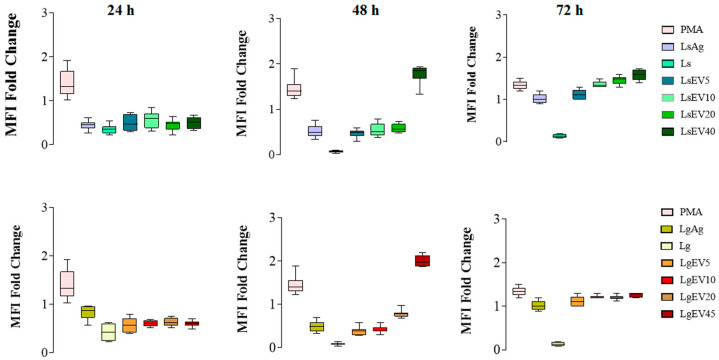
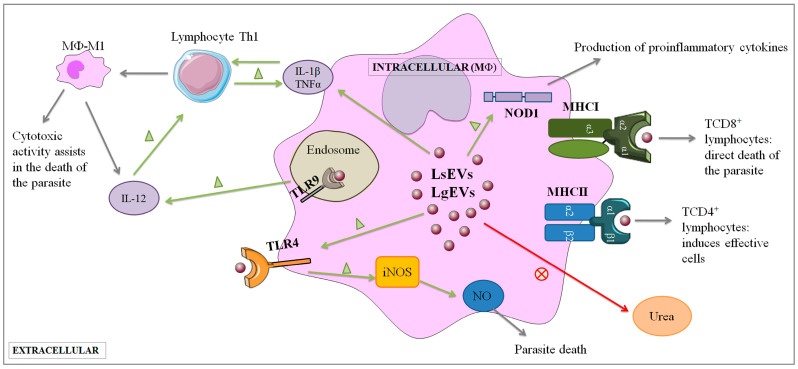
Leishmaniasis is a parasitic disease caused by different species of Leishmania and transmitted through the bite of sand flies vector. Macrophages (MΦ), the target cells of Leishmania parasites, are phagocytes that play a crucial role in the innate immune microbial defense and are antigen-presenting cells driving the activation of the acquired immune response. Exploring parasite-host communication may be key in restraining parasite dissemination in the host. Extracellular vesicles (EVs) constitute a group of heterogenous cell-derived membranous structures, naturally produced by all cells and with immunomodulatory potential over target cells. This study examined the immunogenic potential of EVs shed by L. shawi and L. guyanensis in MΦ activation by analyzing the dynamics of major histocompatibility complex (MHC), innate immune receptors, and cytokine generation. L. shawi and L. guyanensis EVs were incorporated by MΦ and modulated innate immune receptors, indicating that EVs cargo can be recognized by MΦ sensors. Moreover, EVs induced MΦ to generate a mix of pro- and anti-inflammatory cytokines and favored the expression of MHCI molecules, suggesting that EVs antigens can be present to T cells, activating the acquired immune response of the host. Since nano-sized vesicles can be used as vehicles of immune mediators or immunomodulatory drugs, parasitic EVs can be exploited by bioengineering approaches for the development of efficient prophylactic or therapeutic tools for leishmaniasis.
Keywords: Leishmania guyanensis; Leishmania shawi; extracellular vesicles; immunomodulation; leishmaniasis; macrophages.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

Similar articles
-
Modulation of fibroblast behavior by Leishmania: a pathway to understanding disease progression in cutaneous leishmaniasis.BMC Microbiol. 2025 Aug 11;25(1):495. doi: 10.1186/s12866-025-04098-x. BMC Microbiol. 2025. PMID: 40790465 Free PMC article.
-
Swarna Bhasma Induces Antigen-Presenting Abilities of Macrophages and Helps Antigen Experienced CD4+ T Cells to Acquire Th1 Phenotypes Against Leishmania donovani Antigens.Biol Trace Elem Res. 2024 Jan;202(1):210-220. doi: 10.1007/s12011-023-03659-3. Epub 2023 Apr 24. Biol Trace Elem Res. 2024. PMID: 37088826 Free PMC article.
-
Activation of macrophages by extracellular vesicles derived from Babesia-infected red blood cells.Infect Immun. 2025 May 13;93(5):e0033324. doi: 10.1128/iai.00333-24. Epub 2025 Apr 2. Infect Immun. 2025. PMID: 40172538 Free PMC article.
-
Management of urinary stones by experts in stone disease (ESD 2025).Arch Ital Urol Androl. 2025 Jun 30;97(2):14085. doi: 10.4081/aiua.2025.14085. Epub 2025 Jun 30. Arch Ital Urol Androl. 2025. PMID: 40583613 Review.
-
Interventions for Old World cutaneous leishmaniasis.Cochrane Database Syst Rev. 2017 Dec 1;12(12):CD005067. doi: 10.1002/14651858.CD005067.pub5. Cochrane Database Syst Rev. 2017. PMID: 29192424 Free PMC article.
Cited by
-
A Difficult-To-Diagnose Case of American Tegumentary Leishmaniasis.Cureus. 2023 Sep 10;15(9):e44971. doi: 10.7759/cureus.44971. eCollection 2023 Sep. Cureus. 2023. PMID: 37701165 Free PMC article.
-
Extracellular Vesicles Derived From Entamoeba histolytica Have an Immunomodulatory Effect on THP-1 Macrophages.J Parasitol Res. 2024 Oct 29;2024:7325606. doi: 10.1155/2024/7325606. eCollection 2024. J Parasitol Res. 2024. PMID: 39502090 Free PMC article.
-
Emu-miR-10a-5p in Echinococcus multilocularis-derived-extracellular vesicles alleviates airway inflammation in mice with allergic asthma by inhibiting macrophage M2a polarization through LIF-mediated JAK1-STAT3 signaling.Front Immunol. 2025 May 27;16:1577349. doi: 10.3389/fimmu.2025.1577349. eCollection 2025. Front Immunol. 2025. PMID: 40496853 Free PMC article.
-
Unveiling the Interplay Between Dendritic Cells and Natural Killer Cells as Key Players in Leishmania Infection.J Immunol Res. 2025 Feb 10;2025:3176927. doi: 10.1155/jimr/3176927. eCollection 2025. J Immunol Res. 2025. PMID: 39963187 Free PMC article.
-
An in-depth exploration of the multifaceted roles of EVs in the context of pathogenic single-cell microorganisms.Microbiol Mol Biol Rev. 2024 Sep 26;88(3):e0003724. doi: 10.1128/mmbr.00037-24. Epub 2024 Jun 13. Microbiol Mol Biol Rev. 2024. PMID: 38869292 Free PMC article. Review.
References
-
- Calvopina M., Aguirre C., Cevallos W., Castillo A., Abbasi I., Warburg A. Case report: Coinfection of Leishmania guyanensis and human immunodeficiency virus-acquired immune deficiency syndrome: Report of a case of disseminated cutaneous leishmaniasis in Ecuador. Am. J. Trop. Med. Hyg. 2017;96:1151–1154. doi: 10.4269/ajtmh.16-0431. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
