

PARP1 Regulates Circular RNA Biogenesis though Control of Transcriptional Dynamics
- PMID: 37190069
- PMCID: PMC10136798
- DOI: 10.3390/cells12081160
PARP1 Regulates Circular RNA Biogenesis though Control of Transcriptional Dynamics
Abstract
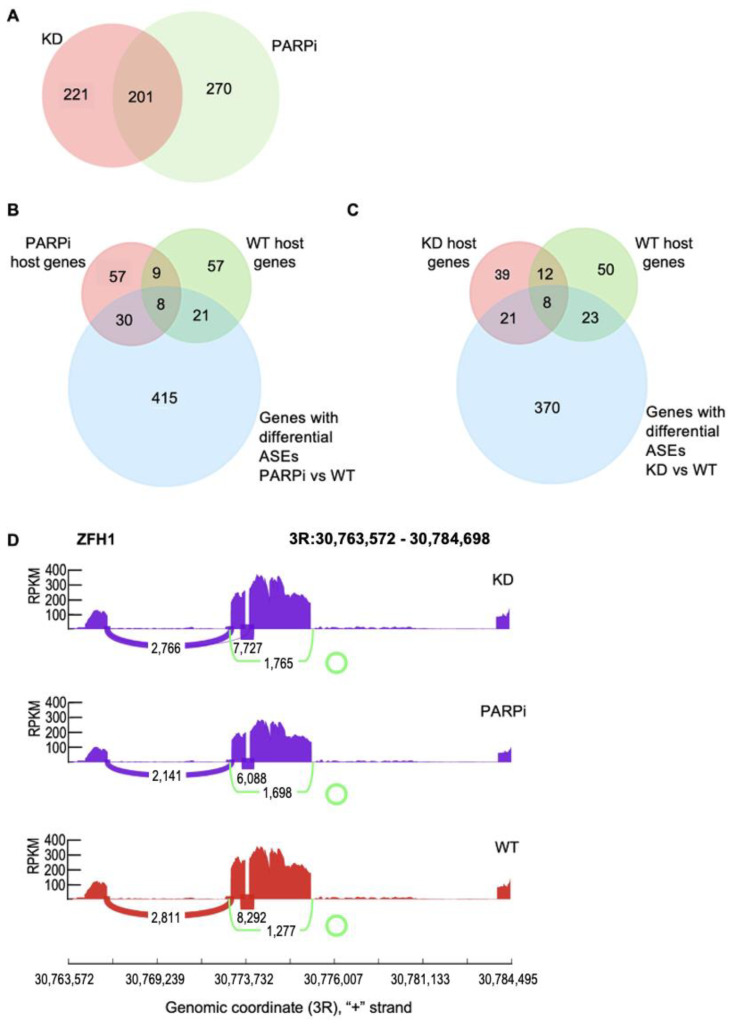
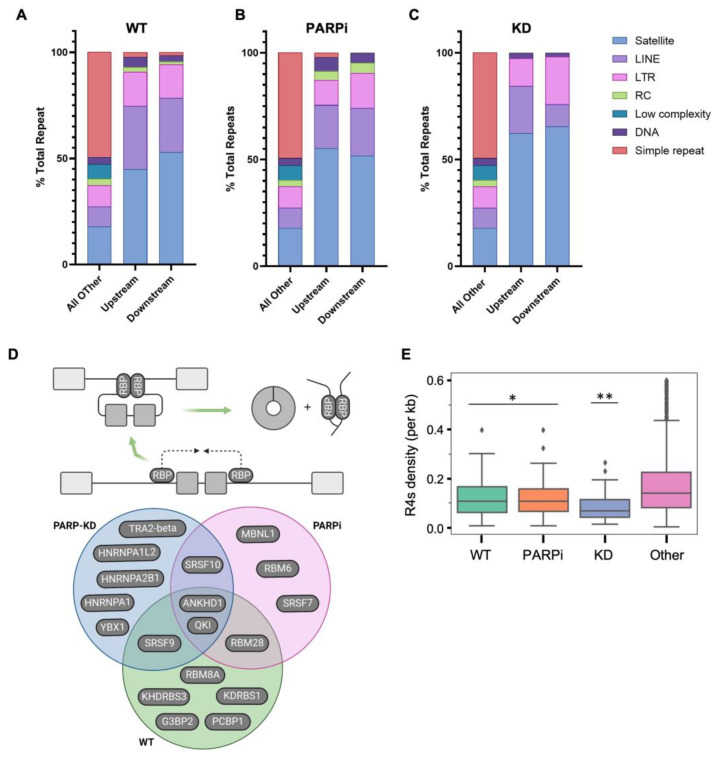
Circular RNAs (circRNAs) are a recently discovered class of RNAs derived from protein-coding genes that have important biological and pathological roles. They are formed through backsplicing during co-transcriptional alternative splicing; however, the unified mechanism that accounts for backsplicing decisions remains unclear. Factors that regulate the transcriptional timing and spatial organization of pre-mRNA, including RNAPII kinetics, the availability of splicing factors, and features of gene architecture, have been shown to influence backsplicing decisions. Poly (ADP-ribose) polymerase I (PARP1) regulates alternative splicing through both its presence on chromatin as well as its PARylation activity. However, no studies have investigated PARP1's possible role in regulating circRNA biogenesis. Here, we hypothesized that PARP1's role in splicing extends to circRNA biogenesis. Our results identify many unique circRNAs in PARP1 depletion and PARylation-inhibited conditions compared to the wild type. We found that while all genes producing circRNAs share gene architecture features common to circRNA host genes, genes producing circRNAs in PARP1 knockdown conditions had longer upstream introns than downstream introns, whereas flanking introns in wild type host genes were symmetrical. Interestingly, we found that the behavior of PARP1 in regulating RNAPII pausing is distinct between these two classes of host genes. We conclude that the PARP1 pausing of RNAPII works within the context of gene architecture to regulate transcriptional kinetics, and therefore circRNA biogenesis. Furthermore, this regulation of PARP1 within host genes acts to fine tune their transcriptional output with implications in gene function.
Keywords: RNA polymerase II elongation and pausing; backsplicing; circRNAs; gene architecture; poly (ADP-ribose) polymerase; splicing; transcription elongation.
Conflict of interest statement
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
Figures






References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
