

PHGDH preserves one-carbon cycle to confer metabolic plasticity in chemoresistant gastric cancer during nutrient stress
- PMID: 37192160
- PMCID: PMC10214193
- DOI: 10.1073/pnas.2217826120
PHGDH preserves one-carbon cycle to confer metabolic plasticity in chemoresistant gastric cancer during nutrient stress
Abstract
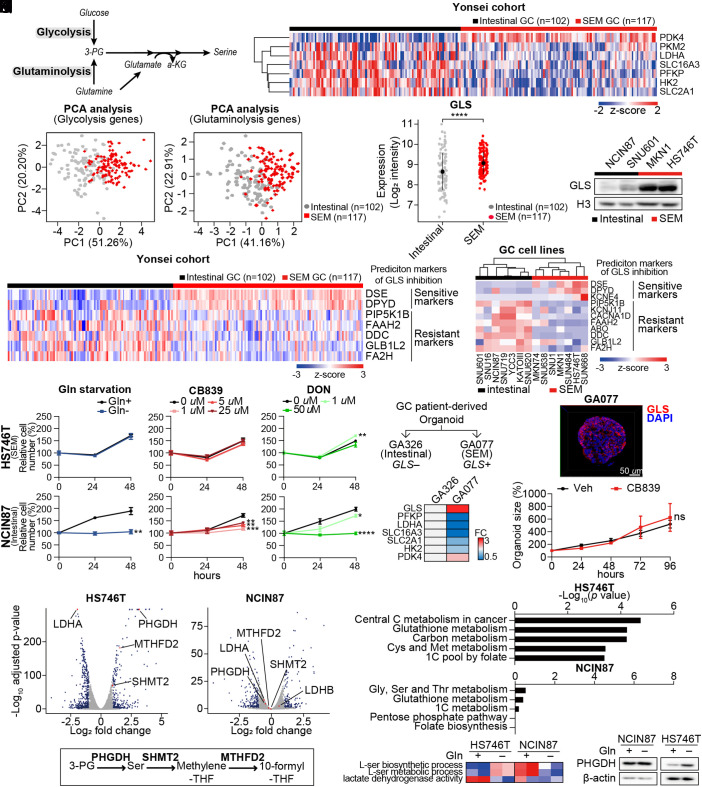
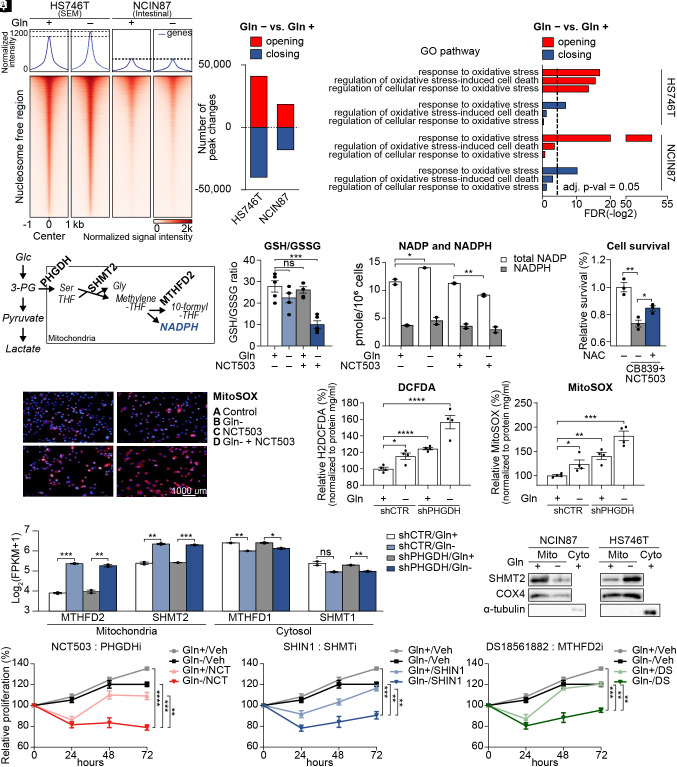
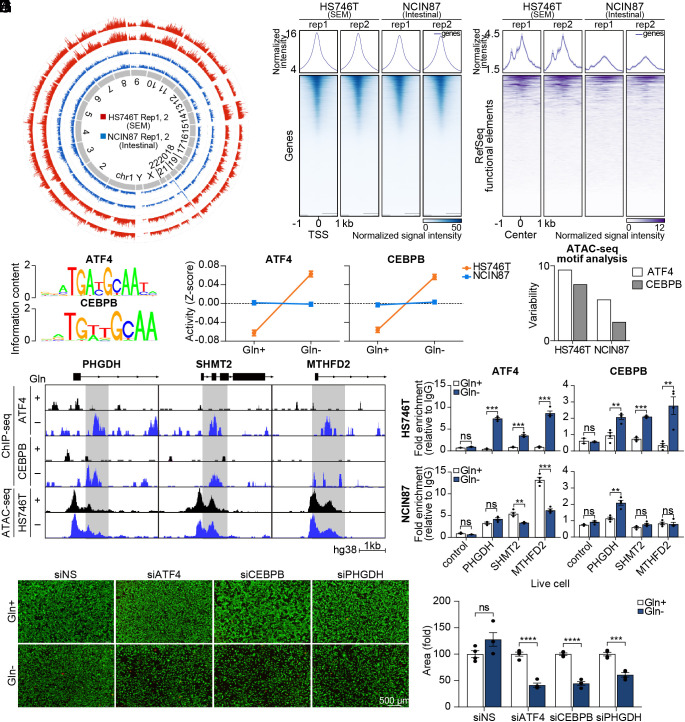
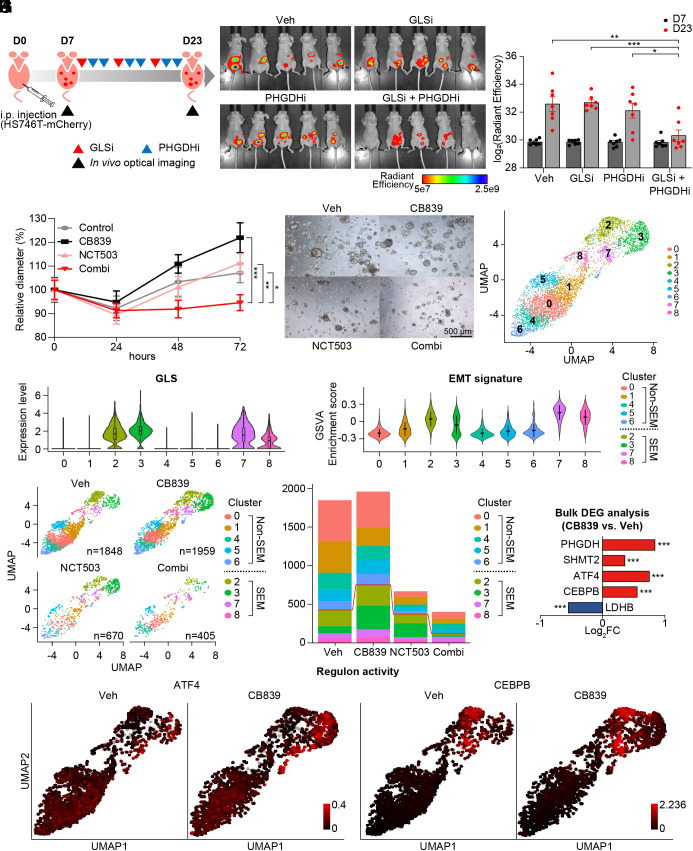
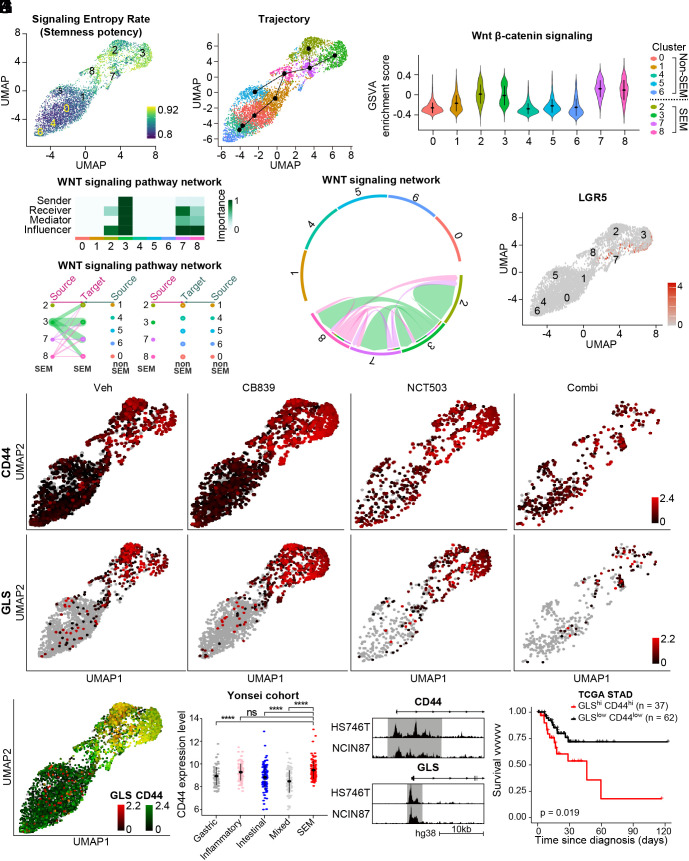
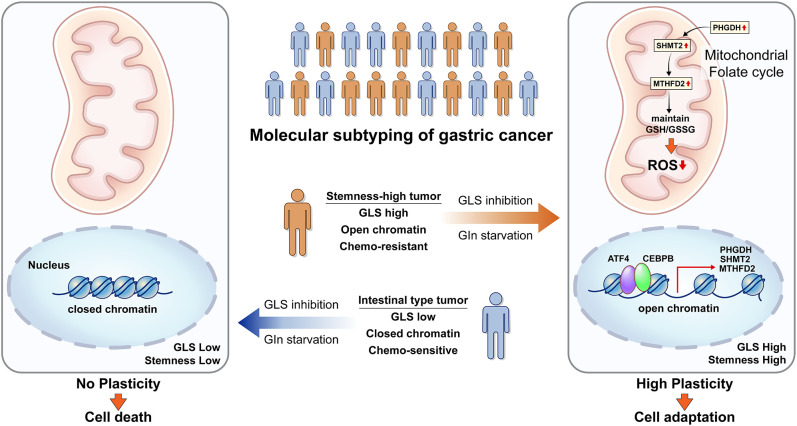
Molecular classification of gastric cancer (GC) identified a subgroup of patients showing chemoresistance and poor prognosis, termed SEM (Stem-like/Epithelial-to-mesenchymal transition/Mesenchymal) type in this study. Here, we show that SEM-type GC exhibits a distinct metabolic profile characterized by high glutaminase (GLS) levels. Unexpectedly, SEM-type GC cells are resistant to glutaminolysis inhibition. We show that under glutamine starvation, SEM-type GC cells up-regulate the 3 phosphoglycerate dehydrogenase (PHGDH)-mediated mitochondrial folate cycle pathway to produce NADPH as a reactive oxygen species scavenger for survival. This metabolic plasticity is associated with globally open chromatin structure in SEM-type GC cells, with ATF4/CEBPB identified as transcriptional drivers of the PHGDH-driven salvage pathway. Single-nucleus transcriptome analysis of patient-derived SEM-type GC organoids revealed intratumoral heterogeneity, with stemness-high subpopulations displaying high GLS expression, a resistance to GLS inhibition, and ATF4/CEBPB activation. Notably, coinhibition of GLS and PHGDH successfully eliminated stemness-high cancer cells. Together, these results provide insight into the metabolic plasticity of aggressive GC cells and suggest a treatment strategy for chemoresistant GC patients.
Keywords: 3 phosphoglycerate dehydrogenase; gastric cancer; glutaminase; metabolic plasticity.
Conflict of interest statement
The authors declare no competing interest.
Figures
References
-
- Cristescu R., et al. , Molecular analysis of gastric cancer identifies subtypes associated with distinct clinical outcomes. Nat. Med. 21, 449–456 (2015). - PubMed
-
- Cheong J. H., et al. , Predictive test for chemotherapy response in resectable gastric cancer: A multi-cohort, retrospective analysis. Lancet Oncol. 19, 629–638 (2018). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
