

GDE7 produces cyclic phosphatidic acid in the ER lumen functioning as a lysophospholipid mediator
- PMID: 37193762
- PMCID: PMC10188492
- DOI: 10.1038/s42003-023-04900-4
GDE7 produces cyclic phosphatidic acid in the ER lumen functioning as a lysophospholipid mediator
Abstract
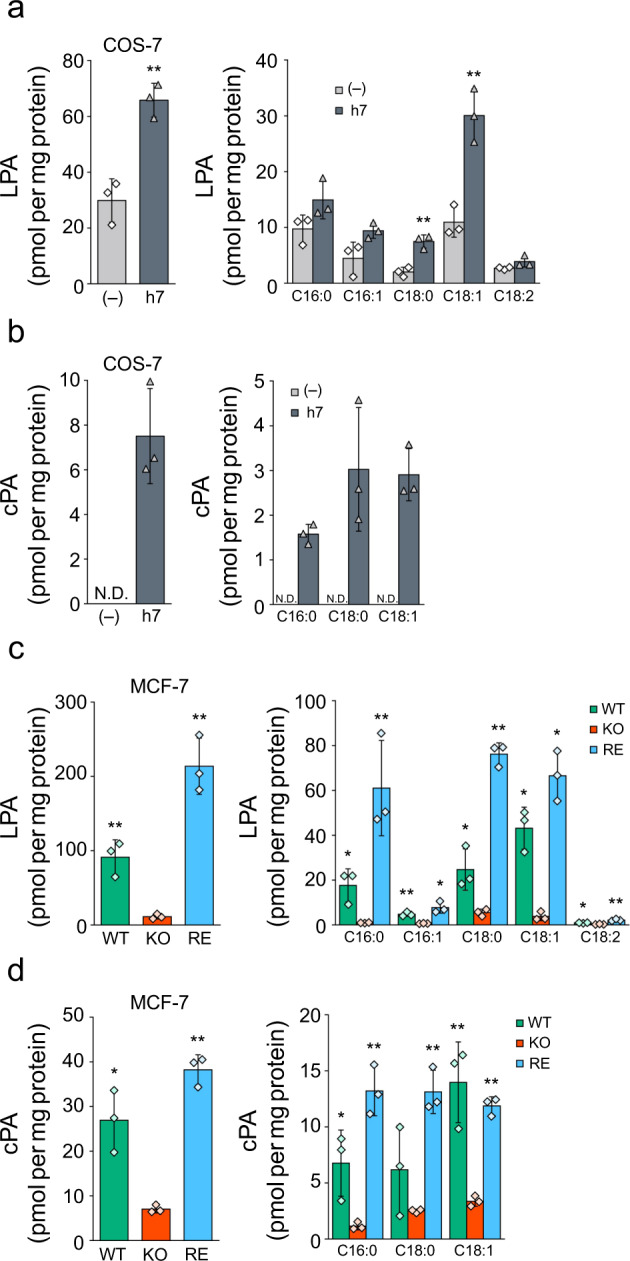
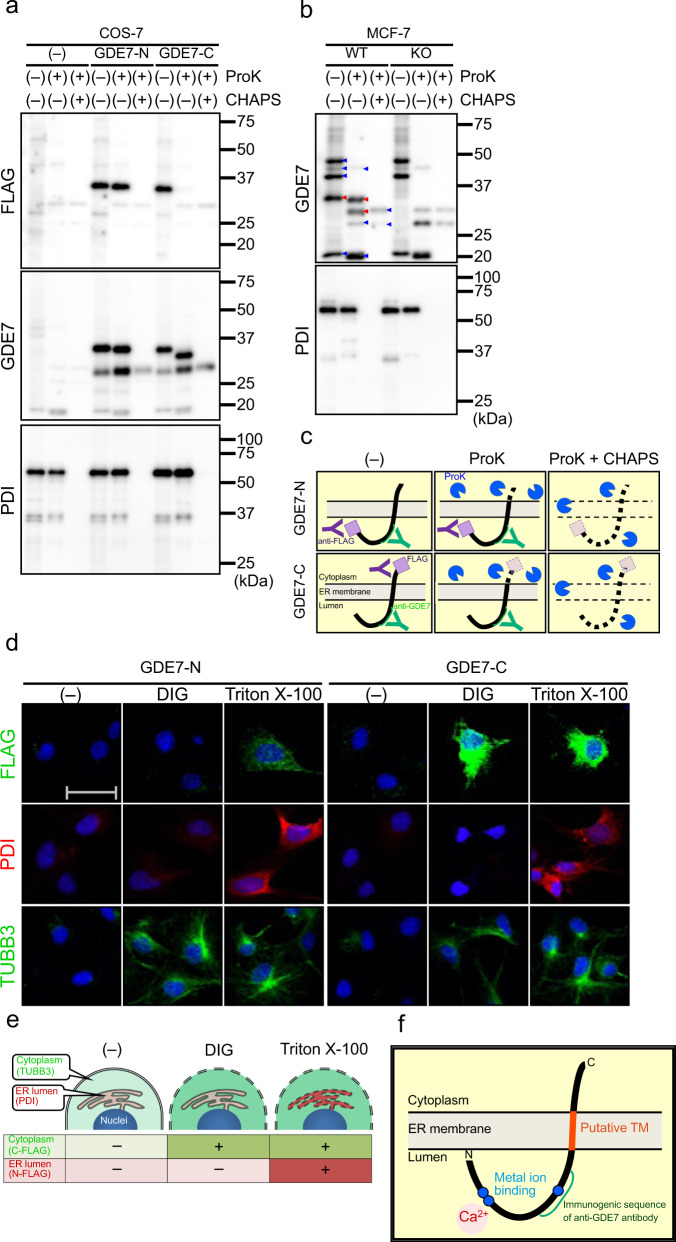
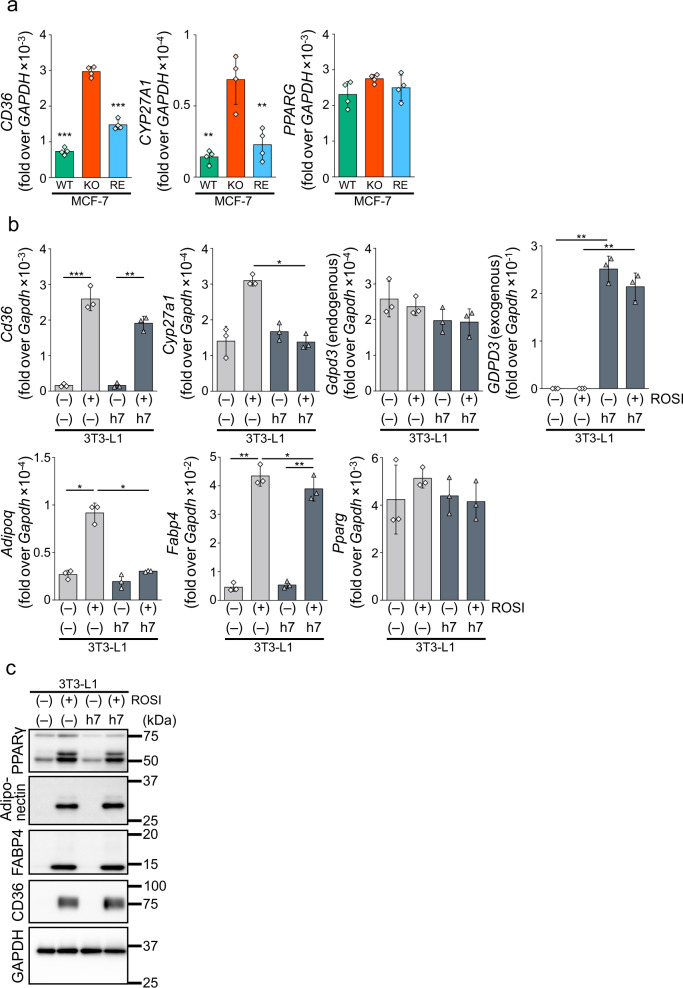
Cyclic phosphatidic acid (cPA) is a lipid mediator, which regulates adipogenic differentiation and glucose homeostasis by suppressing nuclear peroxisome proliferator-activated receptor γ (PPARγ). Glycerophosphodiesterase 7 (GDE7) is a Ca2+-dependent lysophospholipase D that localizes in the endoplasmic reticulum. Although mouse GDE7 catalyzes cPA production in a cell-free system, it is unknown whether GDE7 generates cPA in living cells. Here, we demonstrate that human GDE7 possesses cPA-producing activity in living cells as well as in a cell-free system. Furthermore, the active site of human GDE7 is directed towards the luminal side of the endoplasmic reticulum. Mutagenesis revealed that amino acid residues F227 and Y238 are important for catalytic activity. GDE7 suppresses the PPARγ pathway in human mammary MCF-7 and mouse preadipocyte 3T3-L1 cells, suggesting that cPA functions as an intracellular lipid mediator. These findings lead to a better understanding of the biological role of GDE7 and its product, cPA.
© 2023. The Author(s).
Conflict of interest statement
The authors declare the following competing interests: K.K. and Y.O. received grants from Teijin Pharma Limited, Bayer Yakuhin, Ltd., Kyowa Kirin Co., Ltd., Teijin Nakashima Medical Co., Ltd., Otsuka Pharmaceutical Co., Ltd., Asahi Kasei Pharma Corporation, EA Pharma Co., Ltd., Senko Medical Instrument Mfg. Co., Ltd., and CSL Behring K.K. H.I. is an employee of Maruho Co., Ltd. All other authors declare no conflicts of interest.
Figures




Similar articles
-
Characterization of recombinant murine GDE4 and GDE7, enzymes producing lysophosphatidic acid and/or cyclic phosphatidic acid.J Biochem. 2022 Jan 7;170(6):713-727. doi: 10.1093/jb/mvab091. J Biochem. 2022. PMID: 34523685
-
Development of a selective fluorescence-based enzyme assay for glycerophosphodiesterase family members GDE4 and GDE7.J Lipid Res. 2021;62:100141. doi: 10.1016/j.jlr.2021.100141. Epub 2021 Oct 18. J Lipid Res. 2021. PMID: 34673020 Free PMC article.
-
Calcium-dependent generation of N-acylethanolamines and lysophosphatidic acids by glycerophosphodiesterase GDE7.Biochim Biophys Acta. 2016 Dec;1861(12 Pt A):1881-1892. doi: 10.1016/j.bbalip.2016.09.008. Epub 2016 Sep 13. Biochim Biophys Acta. 2016. PMID: 27637550
-
Lysophospholipid-Related Diseases and PPARγ Signaling Pathway.Int J Mol Sci. 2017 Dec 16;18(12):2730. doi: 10.3390/ijms18122730. Int J Mol Sci. 2017. PMID: 29258184 Free PMC article. Review.
-
Cyclic phosphatidic acid - a unique bioactive phospholipid.Biochim Biophys Acta. 2008 Sep;1781(9):519-24. doi: 10.1016/j.bbalip.2008.05.002. Epub 2008 May 23. Biochim Biophys Acta. 2008. PMID: 18554524 Free PMC article. Review.
Cited by
-
From Classical to Alternative Pathways of 2-Arachidonoylglycerol Synthesis: AlterAGs at the Crossroad of Endocannabinoid and Lysophospholipid Signaling.Molecules. 2024 Aug 4;29(15):3694. doi: 10.3390/molecules29153694. Molecules. 2024. PMID: 39125098 Free PMC article. Review.
-
Recent perspectives on biotechnological production, modulation and applications of glycerophosphoryl diester phosphodiesterases.Biodegradation. 2025 Mar 14;36(2):23. doi: 10.1007/s10532-025-10119-3. Biodegradation. 2025. PMID: 40085296 Review.
-
Characterization of Lysophospholipase D Activity in Mammalian Cell Membranes.Cells. 2024 Mar 16;13(6):520. doi: 10.3390/cells13060520. Cells. 2024. PMID: 38534364 Free PMC article.
-
A Review of Cyclic Phosphatidic Acid and Other Potential Therapeutic Targets for Treating Osteoarthritis.Biomedicines. 2023 Oct 14;11(10):2790. doi: 10.3390/biomedicines11102790. Biomedicines. 2023. PMID: 37893163 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
