

Lysine catabolism reprograms tumour immunity through histone crotonylation
- PMID: 37198486
- PMCID: PMC11089809
- DOI: 10.1038/s41586-023-06061-0
Lysine catabolism reprograms tumour immunity through histone crotonylation
Abstract
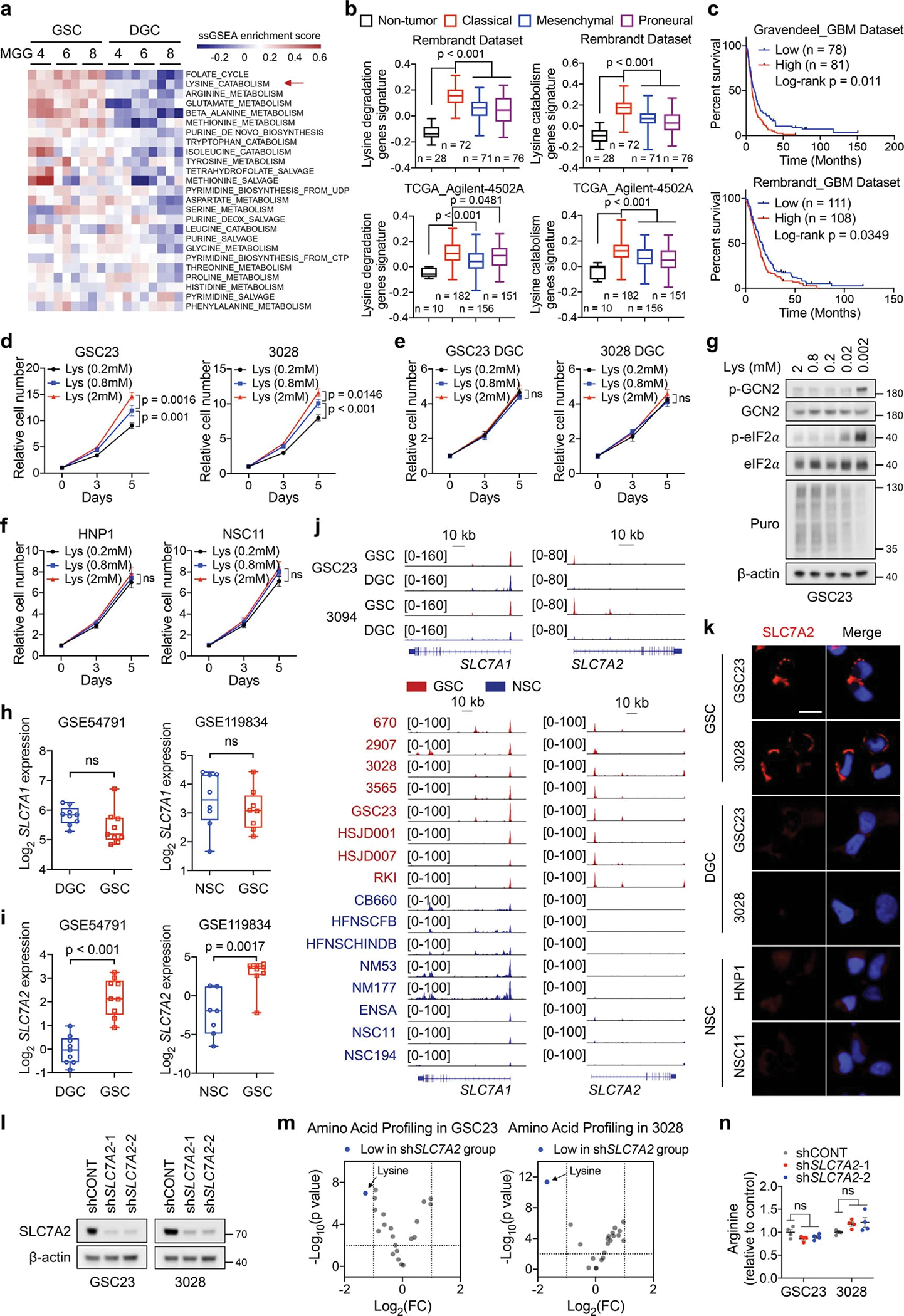
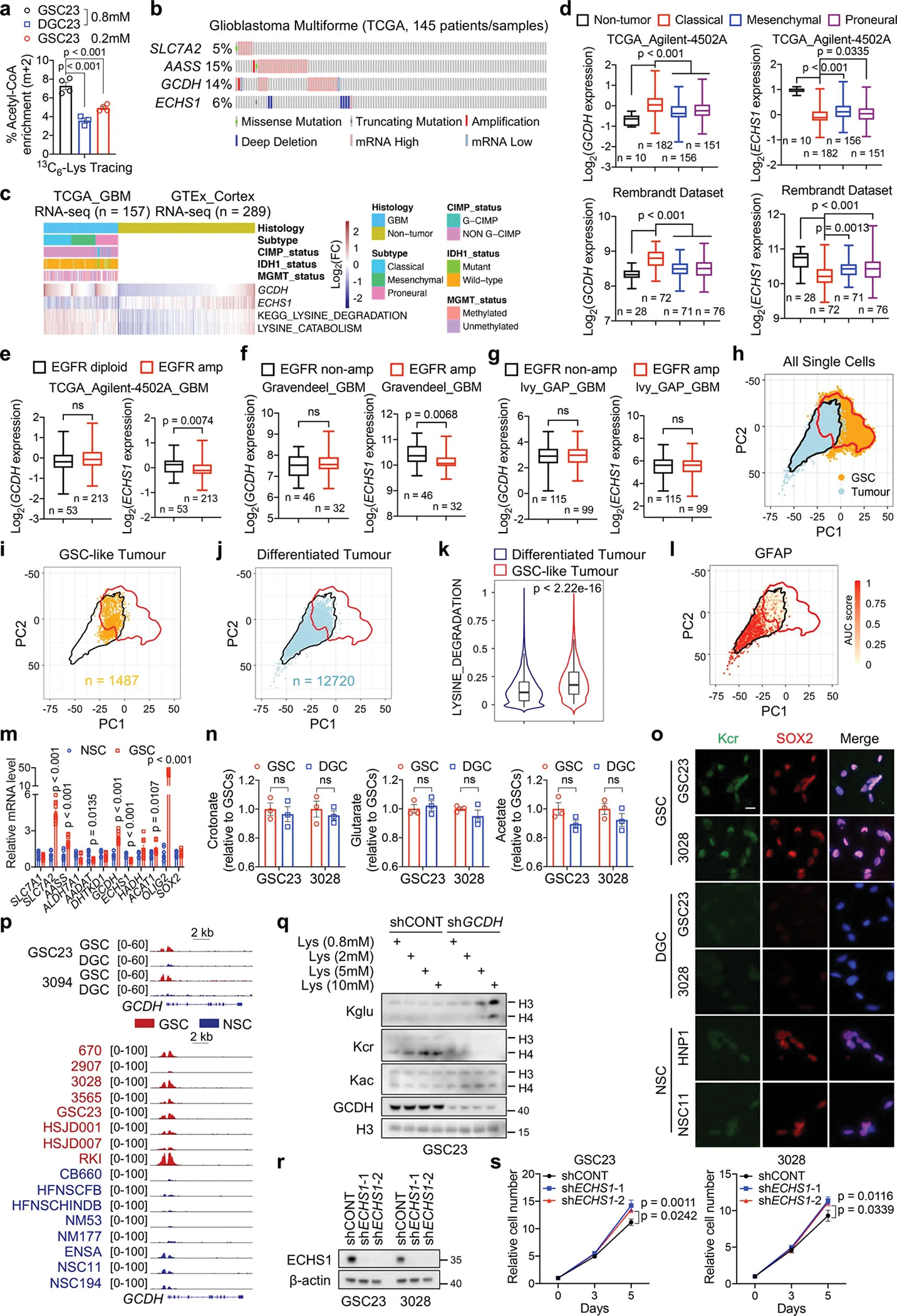
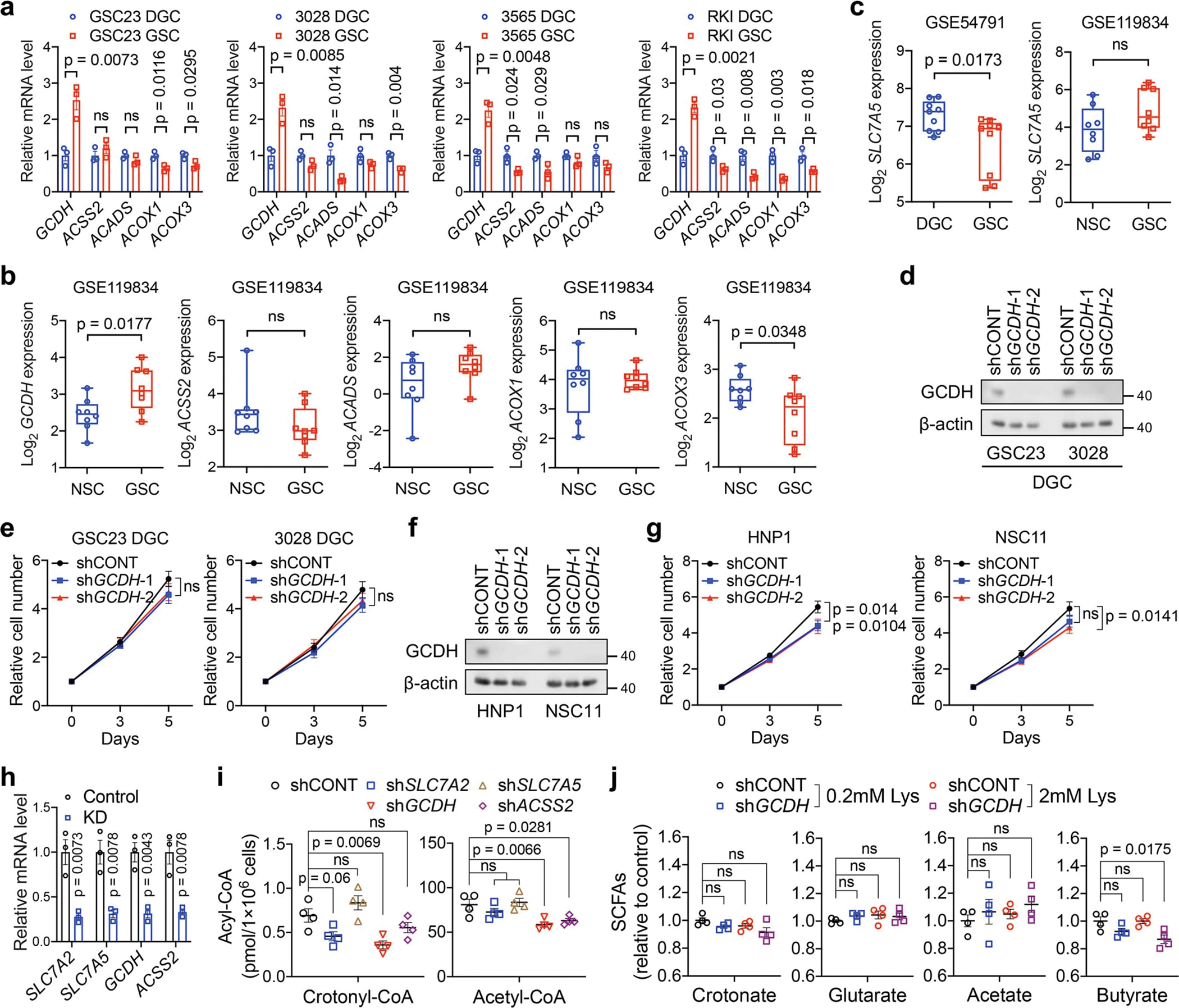
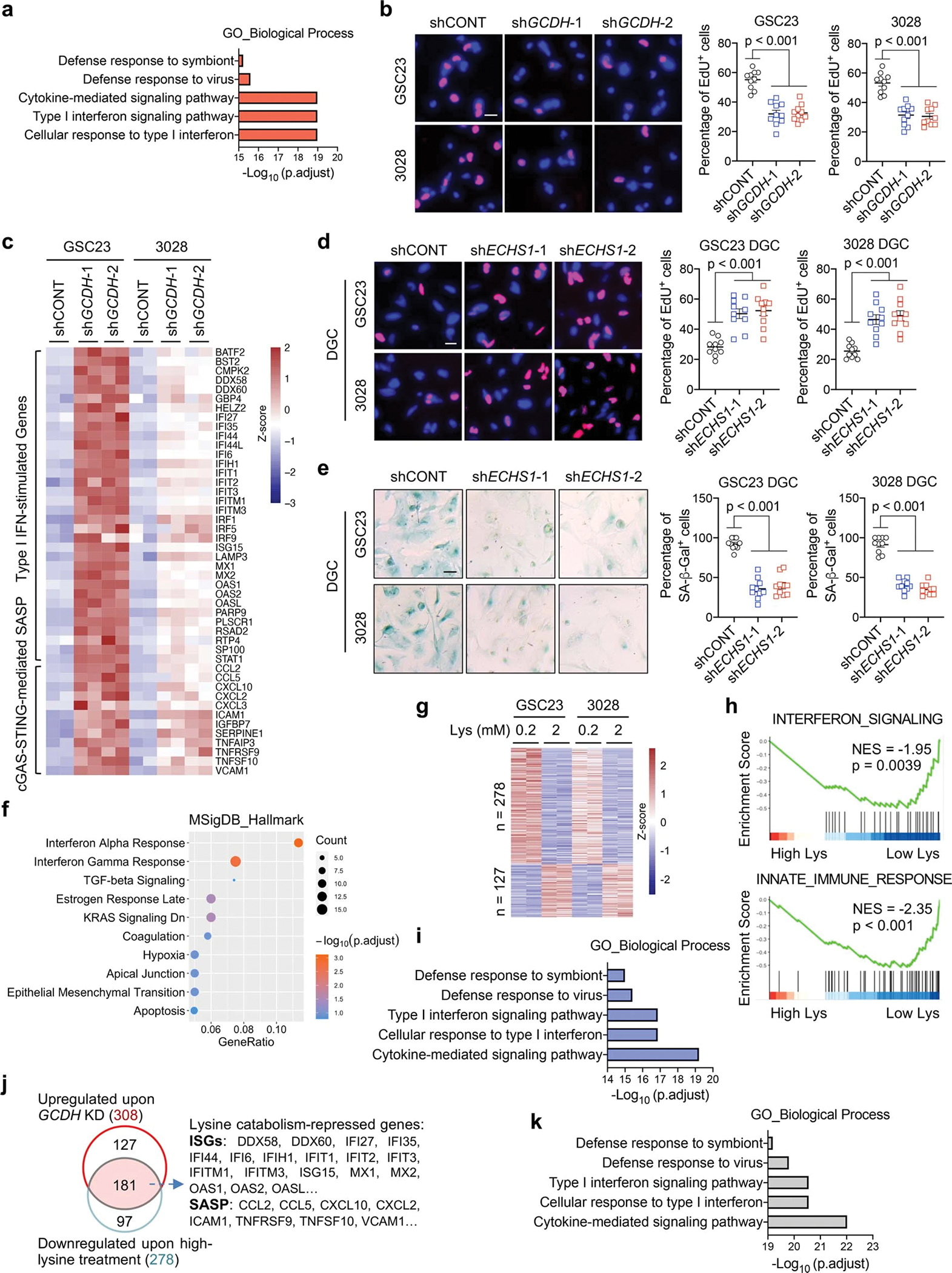
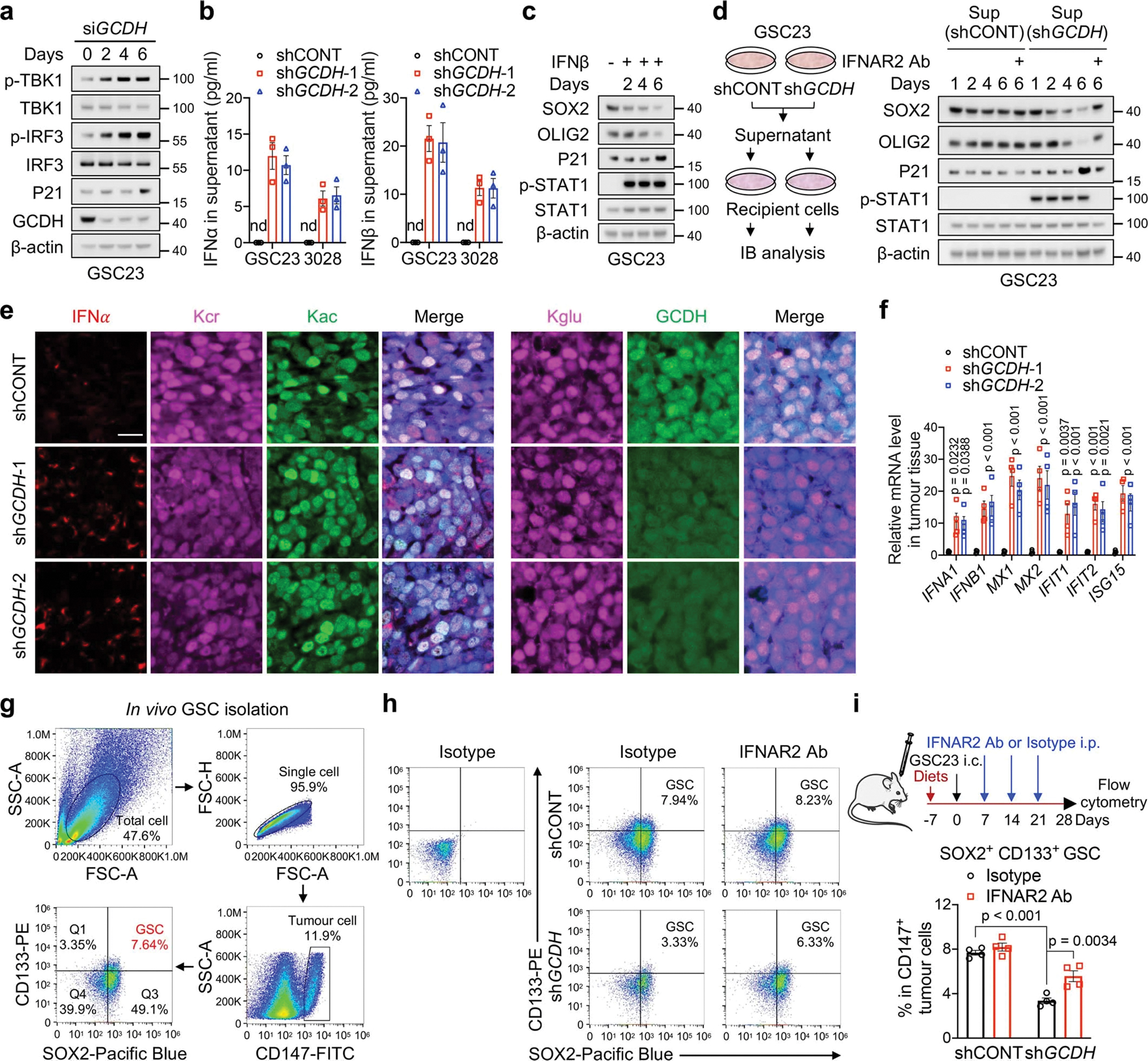
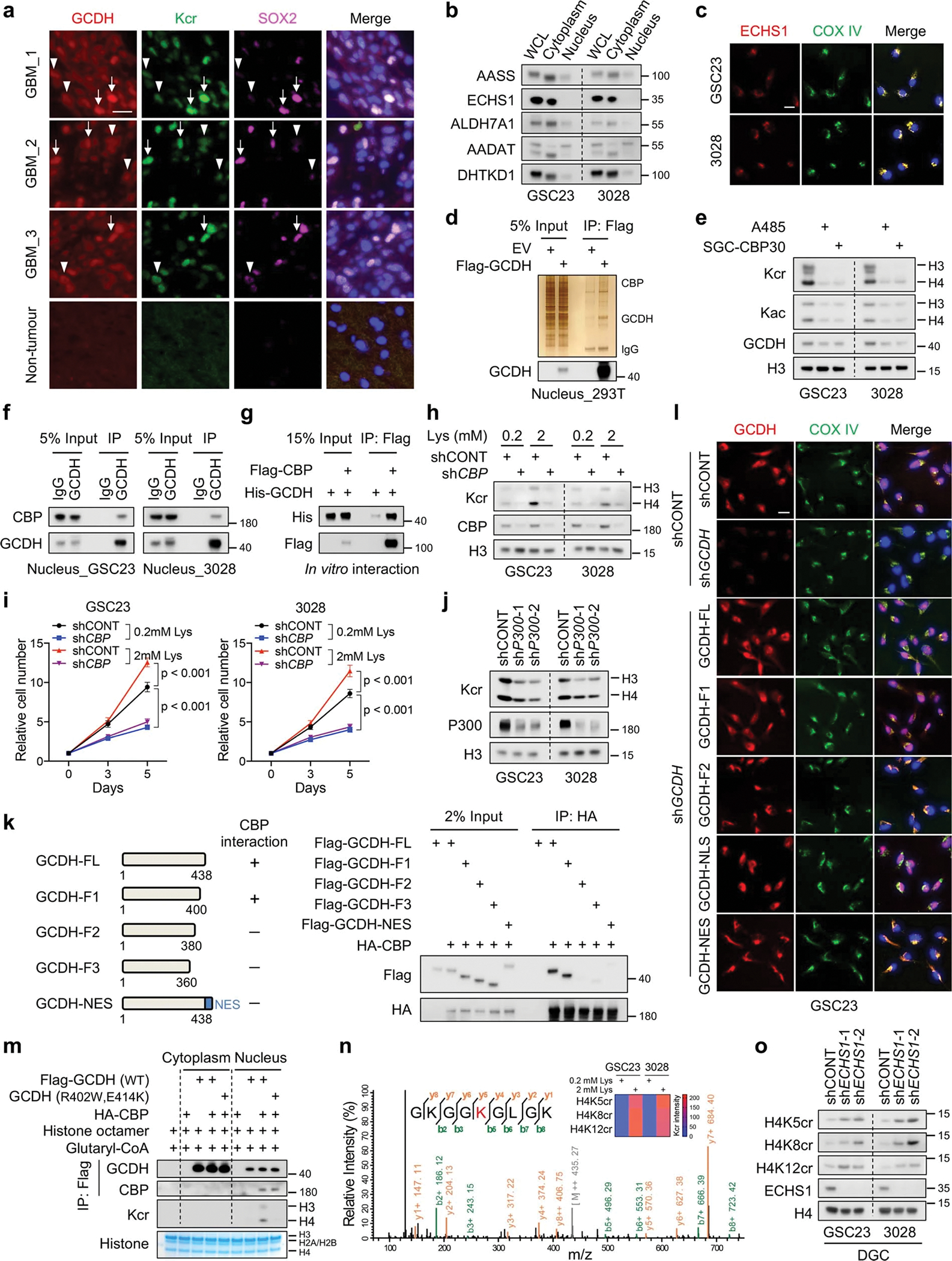
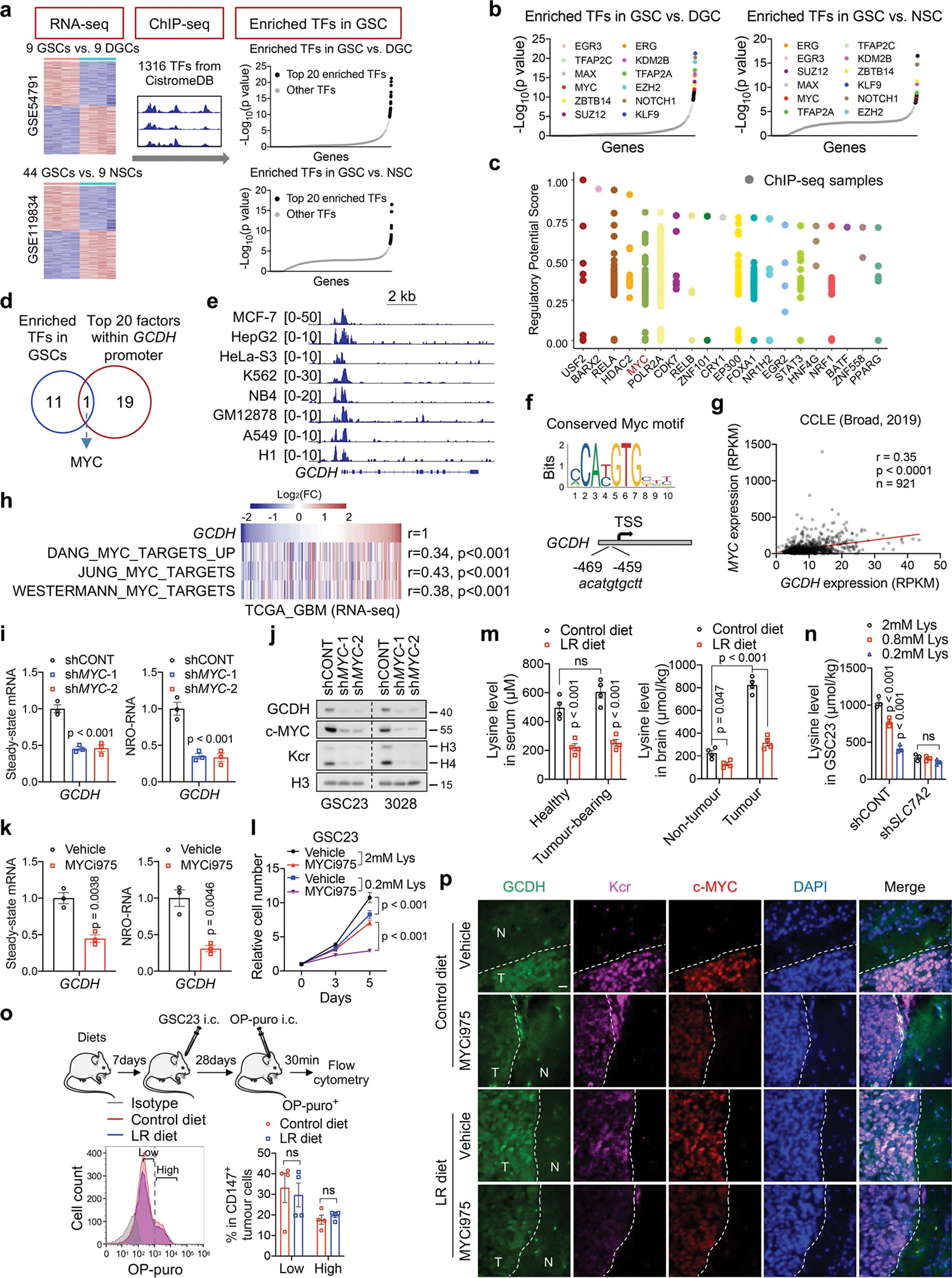
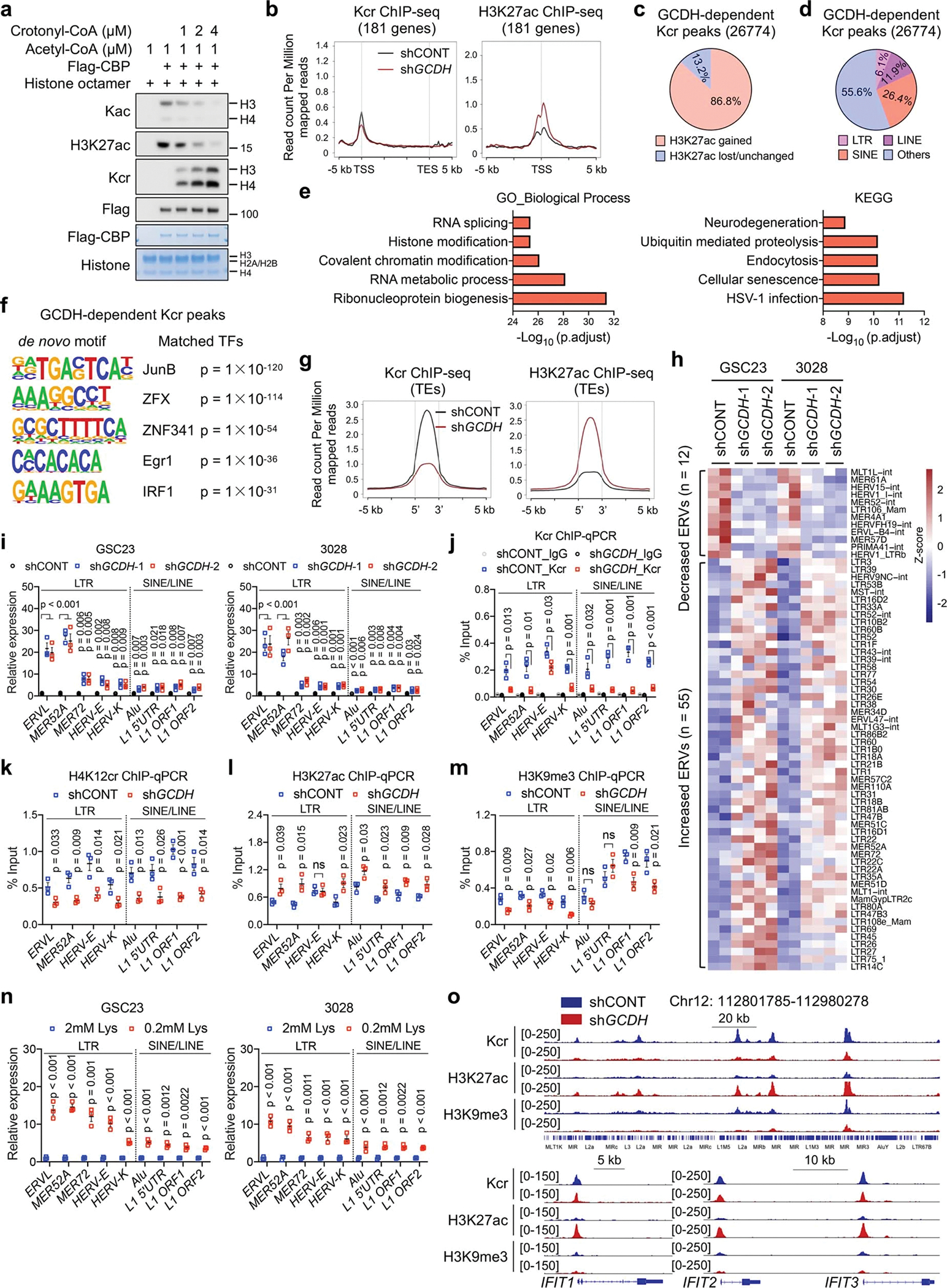
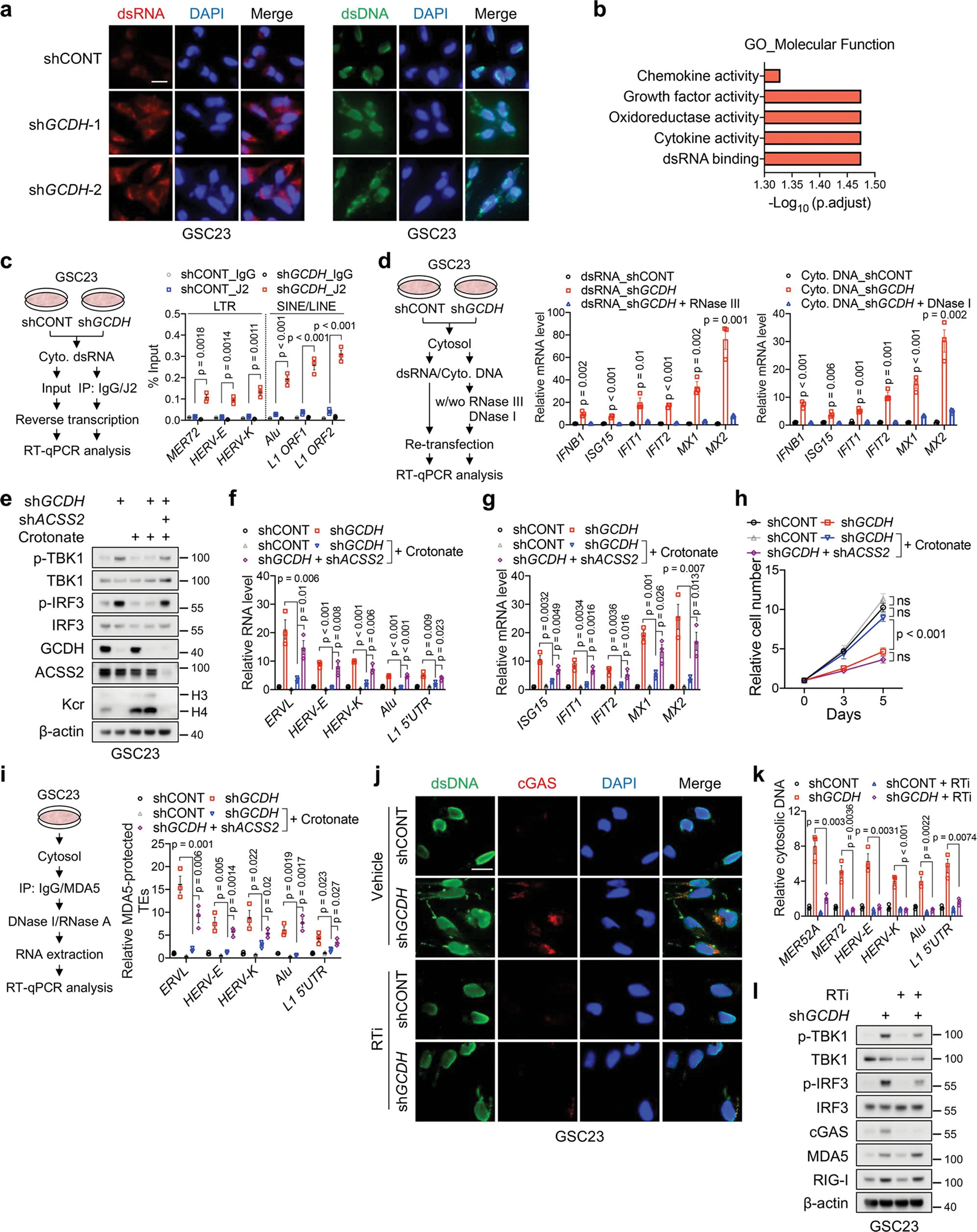
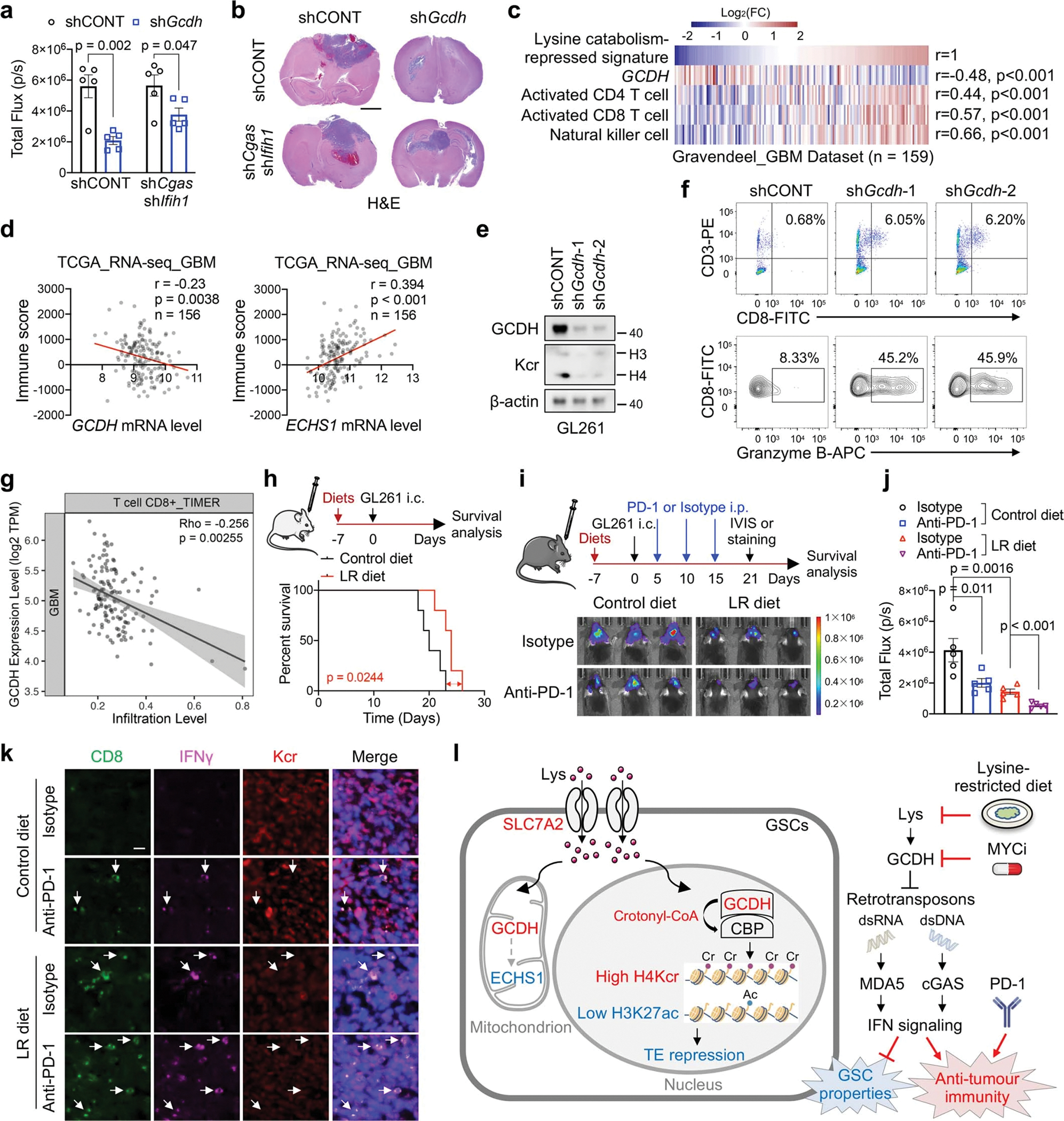
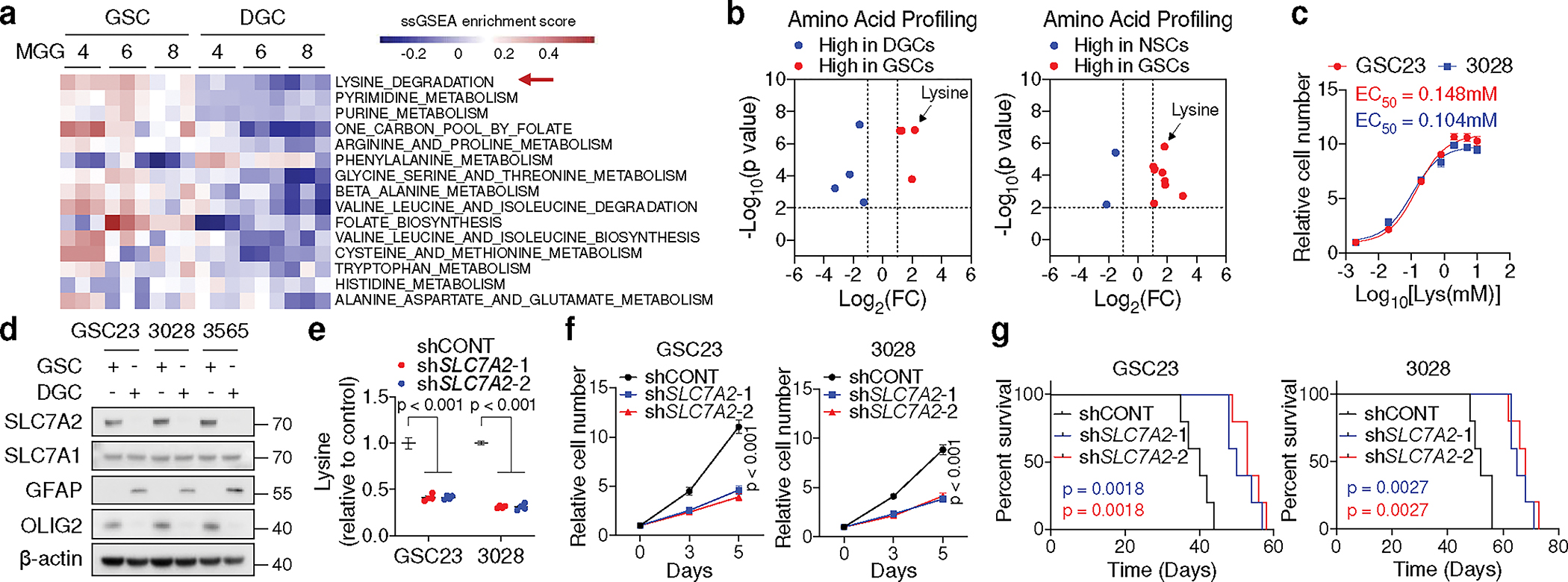
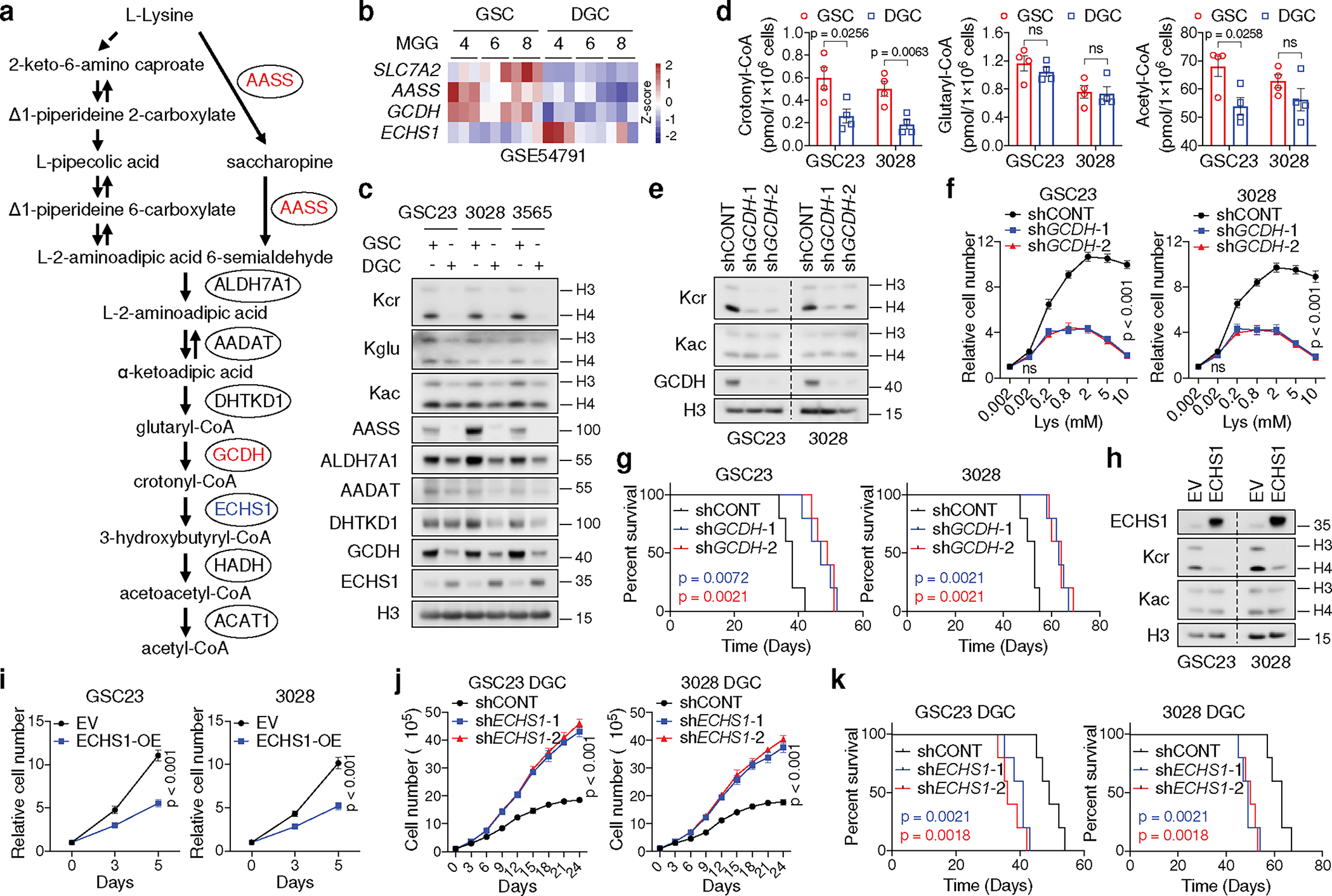
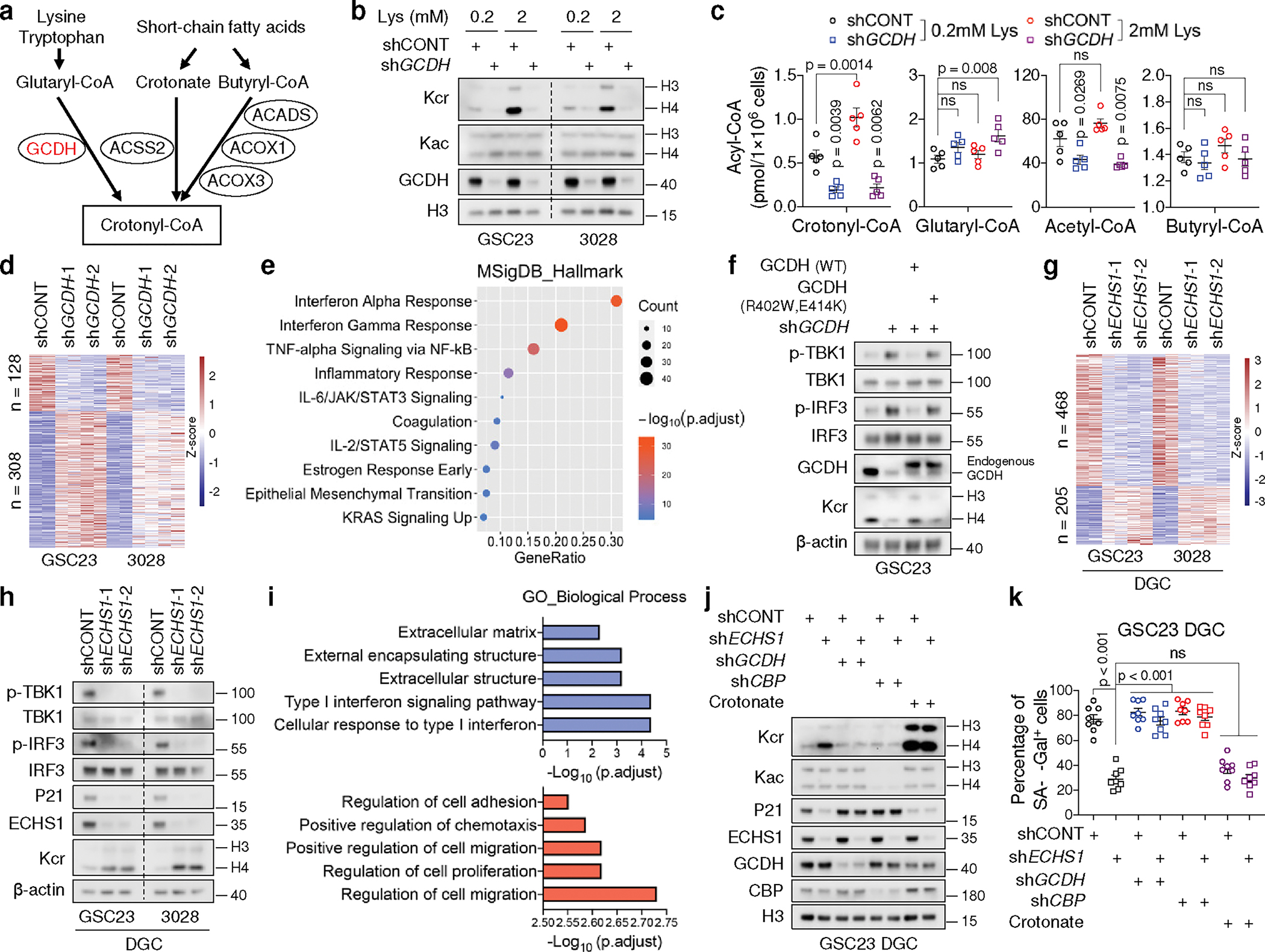
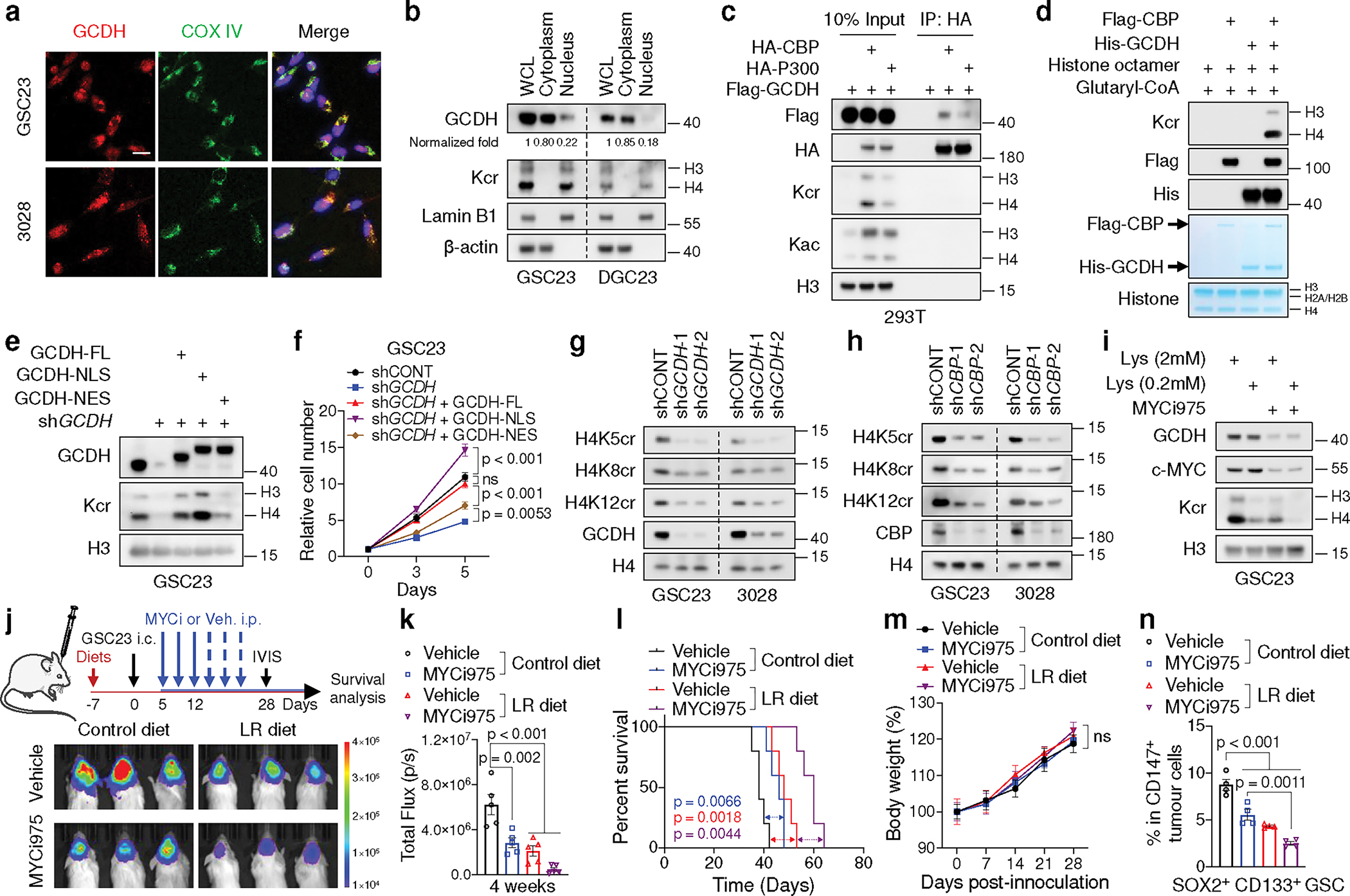
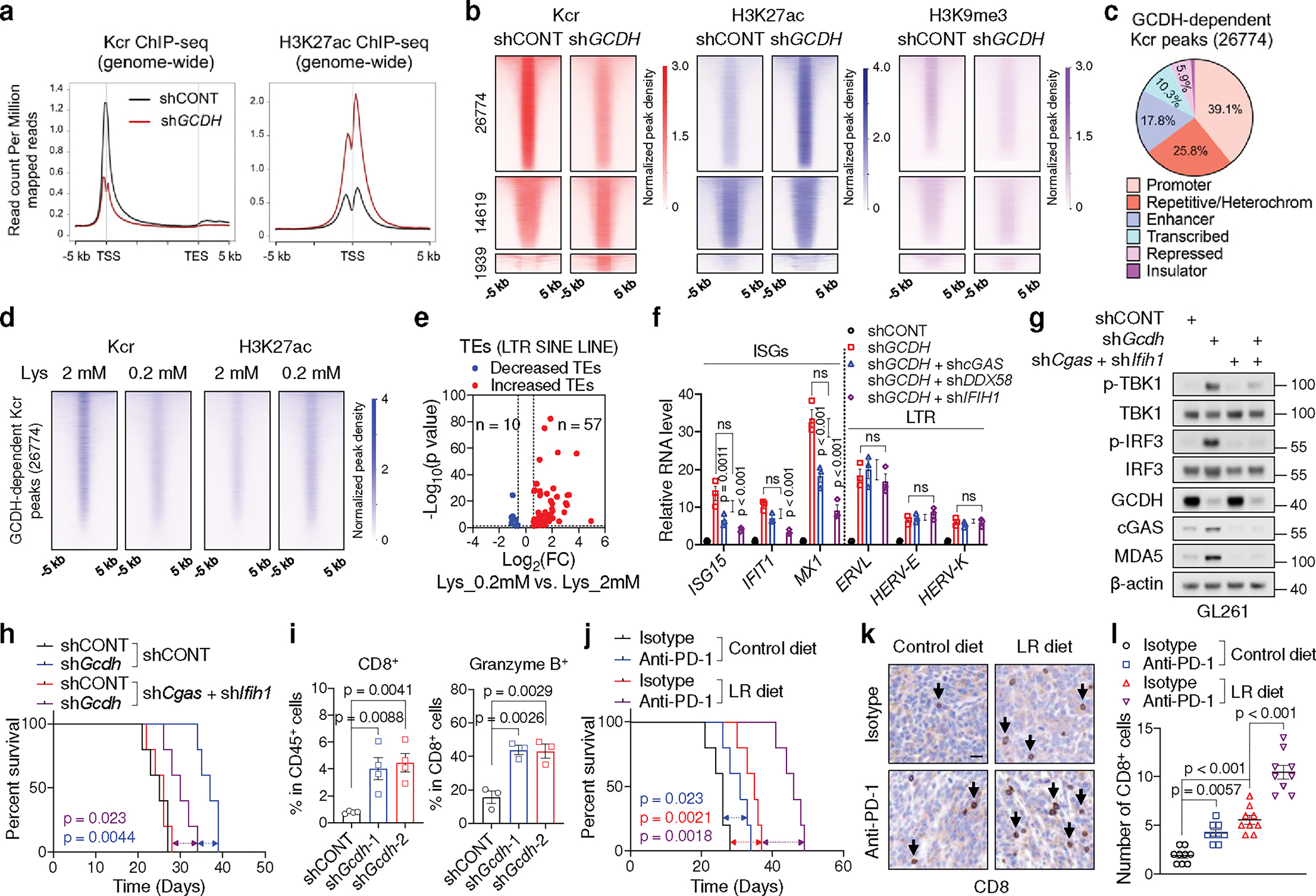
Cancer cells rewire metabolism to favour the generation of specialized metabolites that support tumour growth and reshape the tumour microenvironment1,2. Lysine functions as a biosynthetic molecule, energy source and antioxidant3-5, but little is known about its pathological role in cancer. Here we show that glioblastoma stem cells (GSCs) reprogram lysine catabolism through the upregulation of lysine transporter SLC7A2 and crotonyl-coenzyme A (crotonyl-CoA)-producing enzyme glutaryl-CoA dehydrogenase (GCDH) with downregulation of the crotonyl-CoA hydratase enoyl-CoA hydratase short chain 1 (ECHS1), leading to accumulation of intracellular crotonyl-CoA and histone H4 lysine crotonylation. A reduction in histone lysine crotonylation by either genetic manipulation or lysine restriction impaired tumour growth. In the nucleus, GCDH interacts with the crotonyltransferase CBP to promote histone lysine crotonylation. Loss of histone lysine crotonylation promotes immunogenic cytosolic double-stranded RNA (dsRNA) and dsDNA generation through enhanced H3K27ac, which stimulates the RNA sensor MDA5 and DNA sensor cyclic GMP-AMP synthase (cGAS) to boost type I interferon signalling, leading to compromised GSC tumorigenic potential and elevated CD8+ T cell infiltration. A lysine-restricted diet synergized with MYC inhibition or anti-PD-1 therapy to slow tumour growth. Collectively, GSCs co-opt lysine uptake and degradation to shunt the production of crotonyl-CoA, remodelling the chromatin landscape to evade interferon-induced intrinsic effects on GSC maintenance and extrinsic effects on immune response.
© 2023. The Author(s), under exclusive licence to Springer Nature Limited.
Conflict of interest statement
Competing interests
Y.Z. is a founder, board member, advisor to, and inventor on patents licensed to PTM Bio Inc. (Hangzhou, China and Chicago, IL) and Maponos Therapeutics Inc. (Chicago, IL). The other authors declare no competing interests.
Figures
Comment in
-
Lysine metabolism at the nexus of crotonylation and tumor immunity.Cell Res. 2023 Nov;33(11):813-814. doi: 10.1038/s41422-023-00841-z. Cell Res. 2023. PMID: 37380809 Free PMC article. No abstract available.
References
-
- Gimple RC, Yang K, Halbert ME, Agnihotri S & Rich JN Brain cancer stem cells: resilience through adaptive plasticity and hierarchical heterogeneity. Nat. Rev. Cancer 22, 497–514 (2022). - PubMed
-
- Wu Z et al. TPO-Induced Metabolic Reprogramming Drives Liver Metastasis of Colorectal Cancer CD110+ Tumor-Initiating Cells. Cell Stem Cell 17, 47–59 (2015). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
