

RNA conformational propensities determine cellular activity
- PMID: 37198487
- PMCID: PMC10429349
- DOI: 10.1038/s41586-023-06080-x
RNA conformational propensities determine cellular activity
Erratum in
-
Author Correction: RNA conformational propensities determine cellular activity.Nature. 2023 Jun;618(7967):E35. doi: 10.1038/s41586-023-06286-z. Nature. 2023. PMID: 37296211 No abstract available.
Abstract
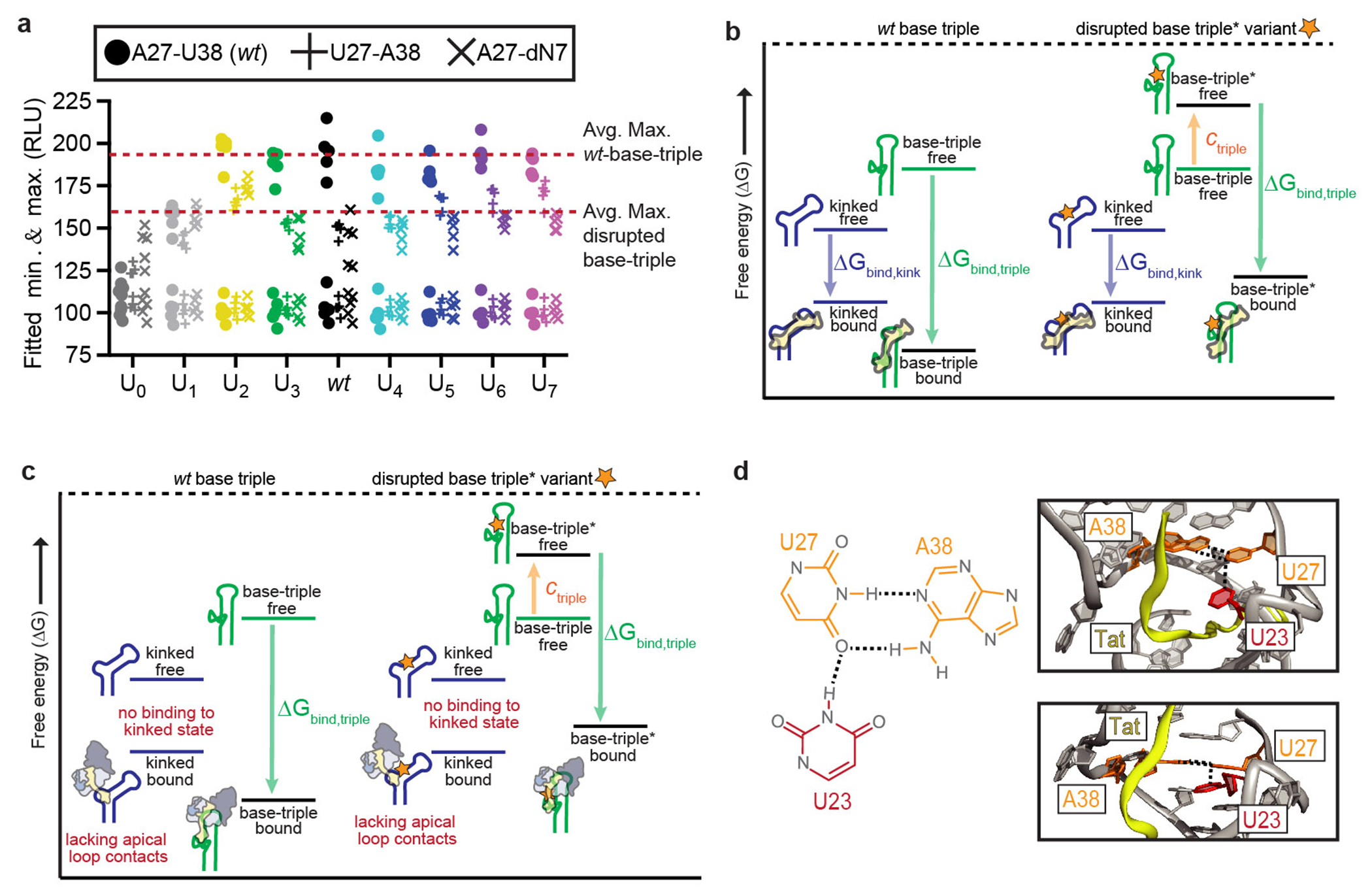
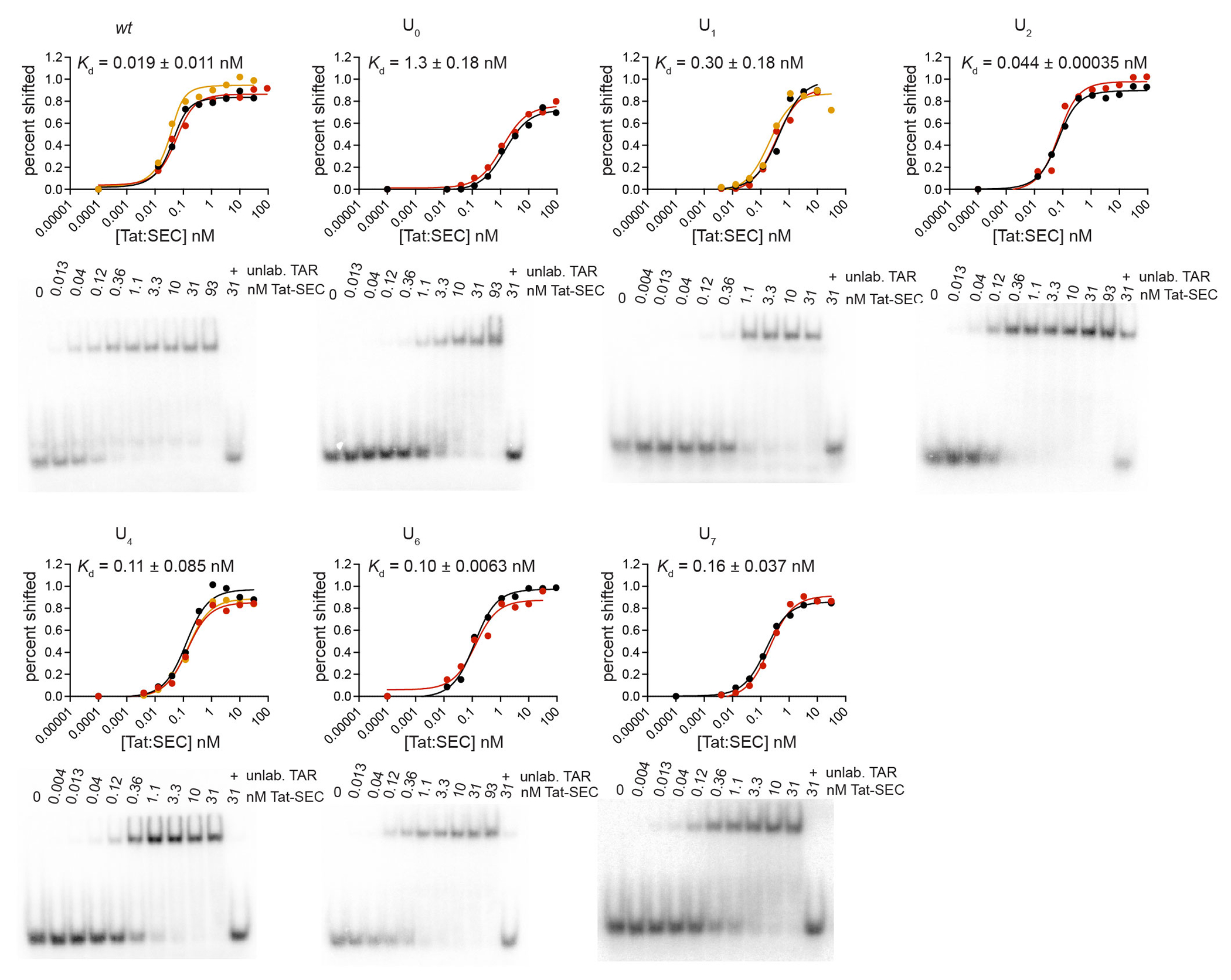
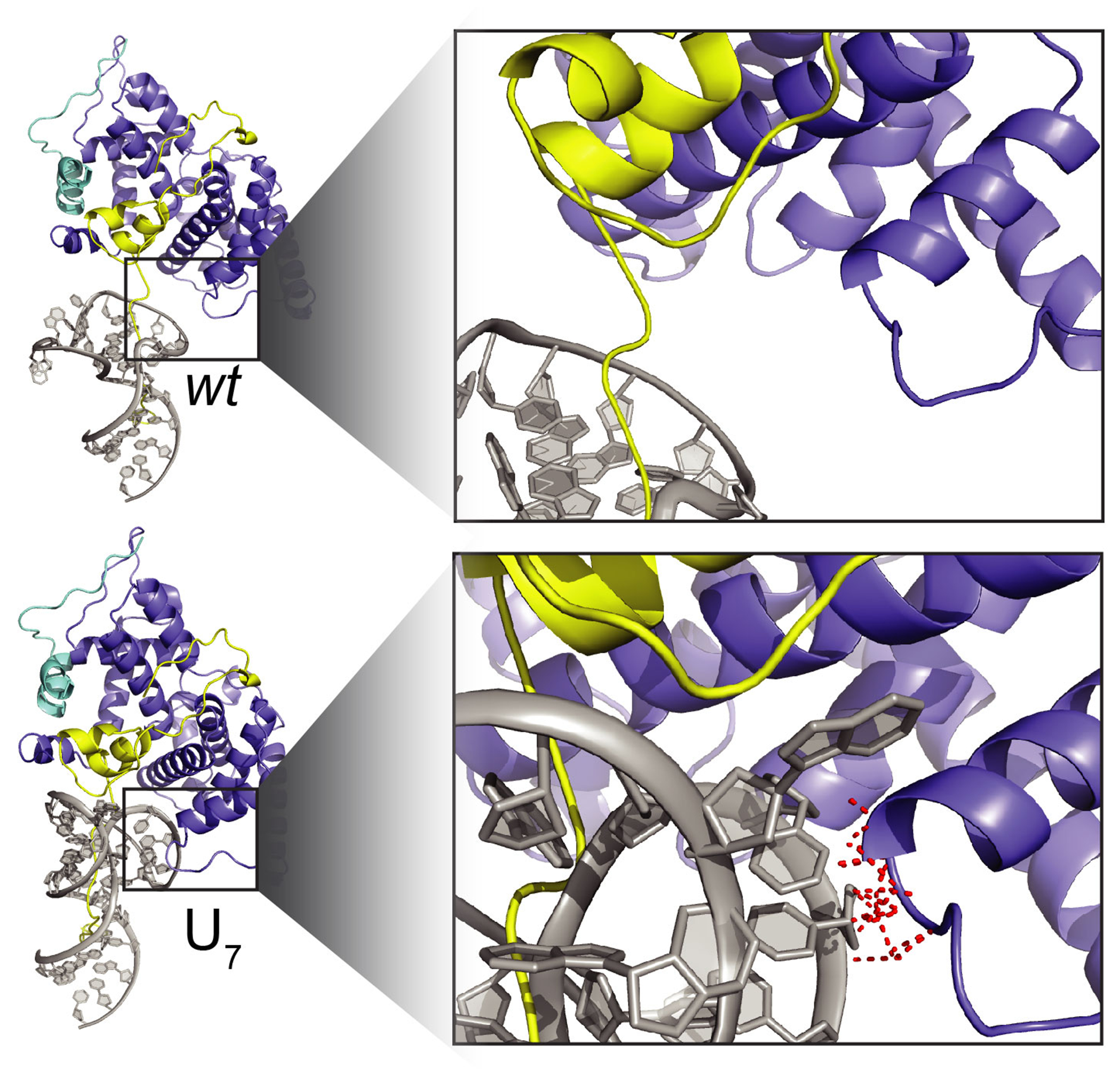
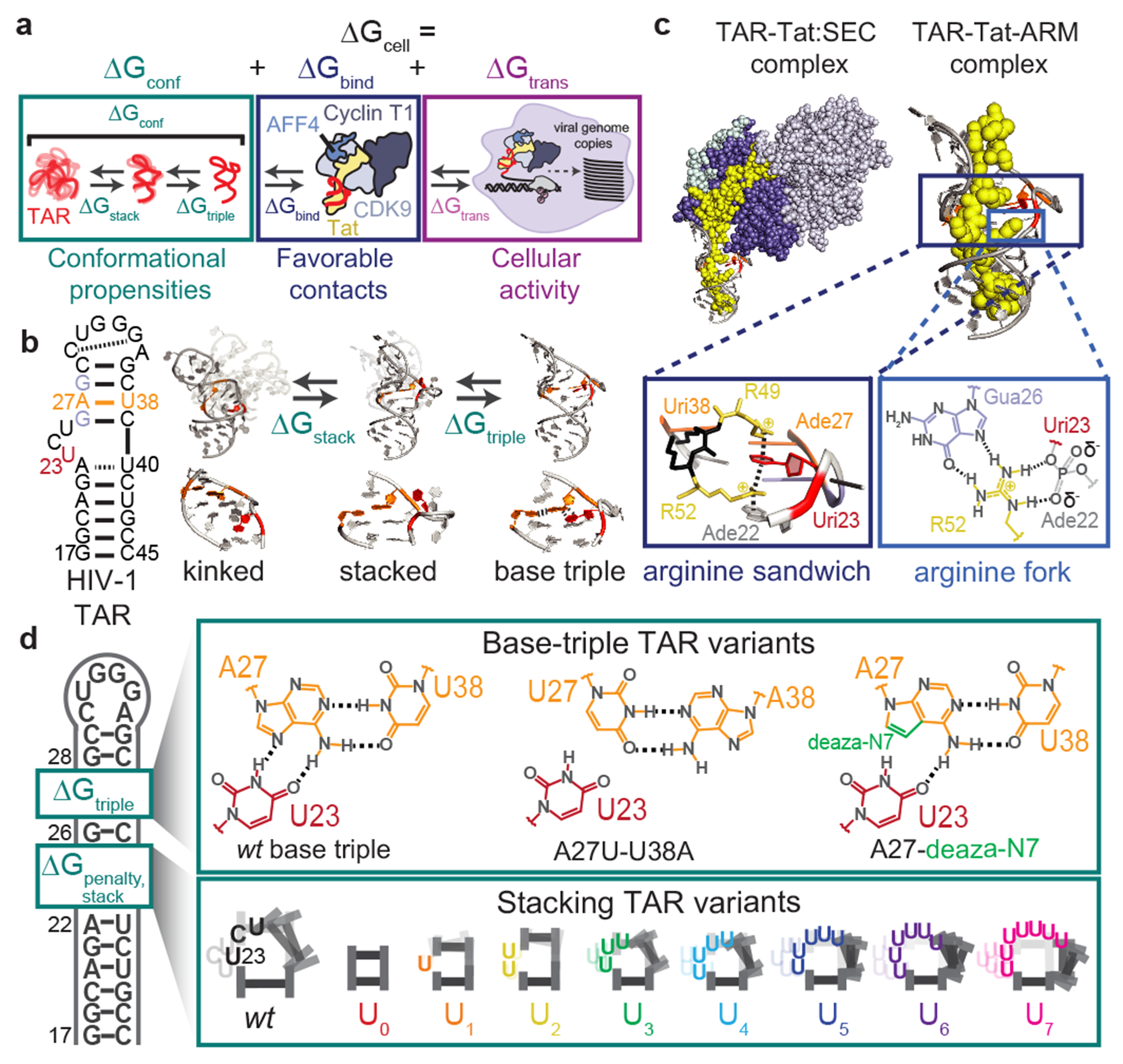
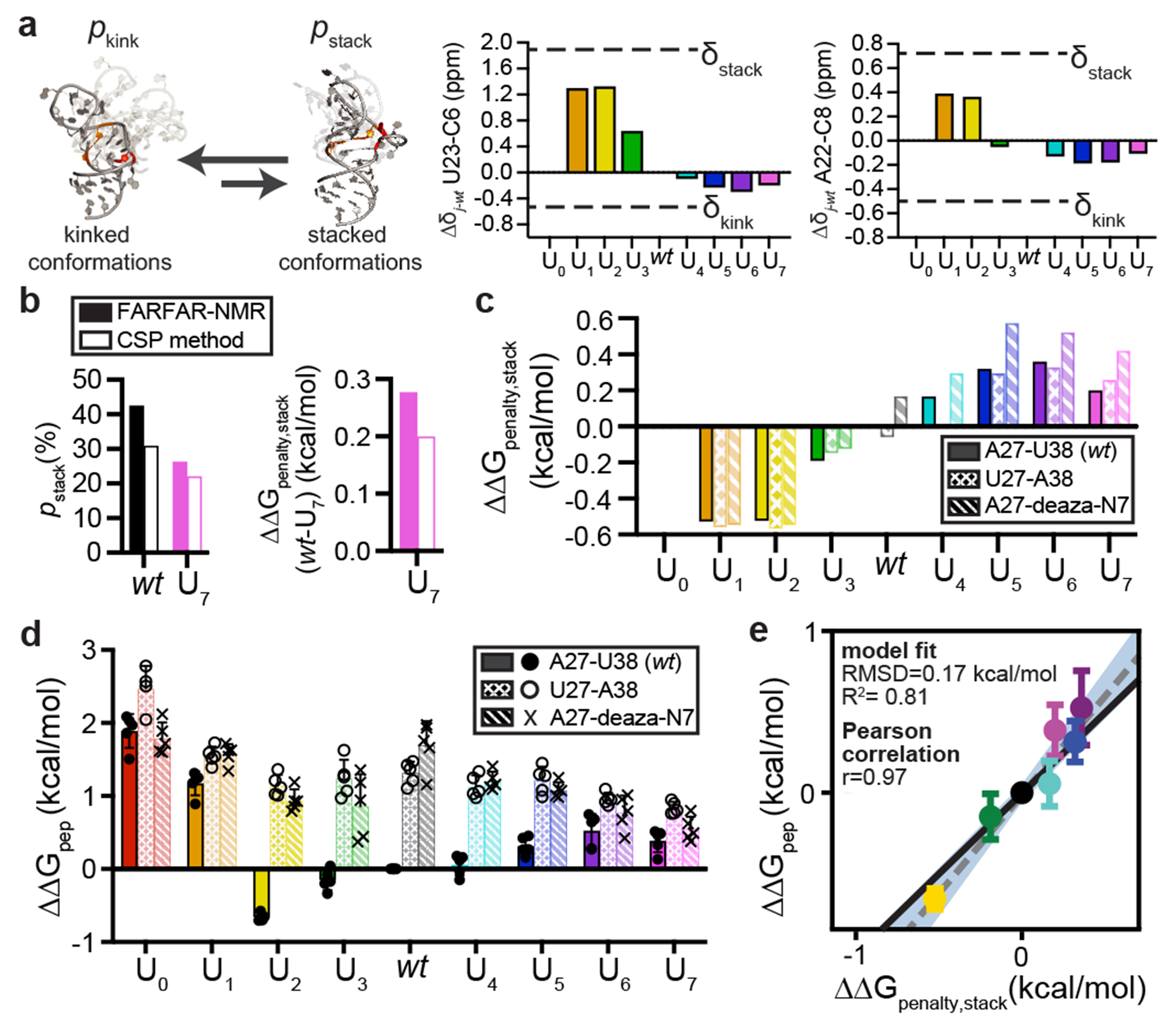
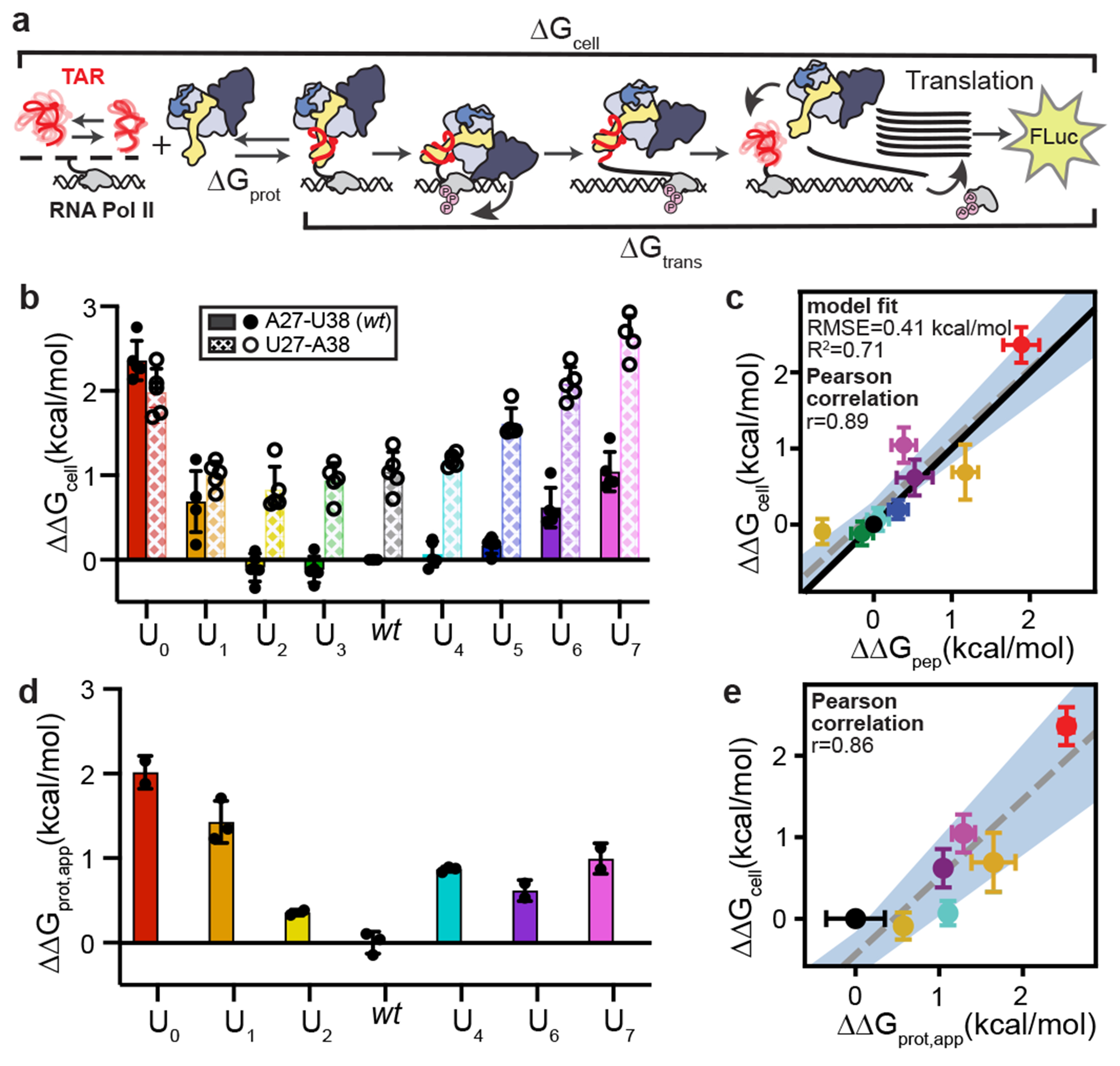
Cellular processes are the product of interactions between biomolecules, which associate to form biologically active complexes1. These interactions are mediated by intermolecular contacts, which if disrupted, lead to alterations in cell physiology. Nevertheless, the formation of intermolecular contacts nearly universally requires changes in the conformations of the interacting biomolecules. As a result, binding affinity and cellular activity crucially depend both on the strength of the contacts and on the inherent propensities to form binding-competent conformational states2,3. Thus, conformational penalties are ubiquitous in biology and must be known in order to quantitatively model binding energetics for protein and nucleic acid interactions4,5. However, conceptual and technological limitations have hindered our ability to dissect and quantitatively measure how conformational propensities affect cellular activity. Here we systematically altered and determined the propensities for forming the protein-bound conformation of HIV-1 TAR RNA. These propensities quantitatively predicted the binding affinities of TAR to the RNA-binding region of the Tat protein and predicted the extent of HIV-1 Tat-dependent transactivation in cells. Our results establish the role of ensemble-based conformational propensities in cellular activity and reveal an example of a cellular process driven by an exceptionally rare and short-lived RNA conformational state.
© 2023. The Author(s), under exclusive licence to Springer Nature Limited.
Conflict of interest statement
Competing interest declaration
HMA is an adviser to and holds an ownership interest in Base4, an RNA-based drug discovery company. DH is a consultant for Radial, an RNA-based drug discovery company.
Figures

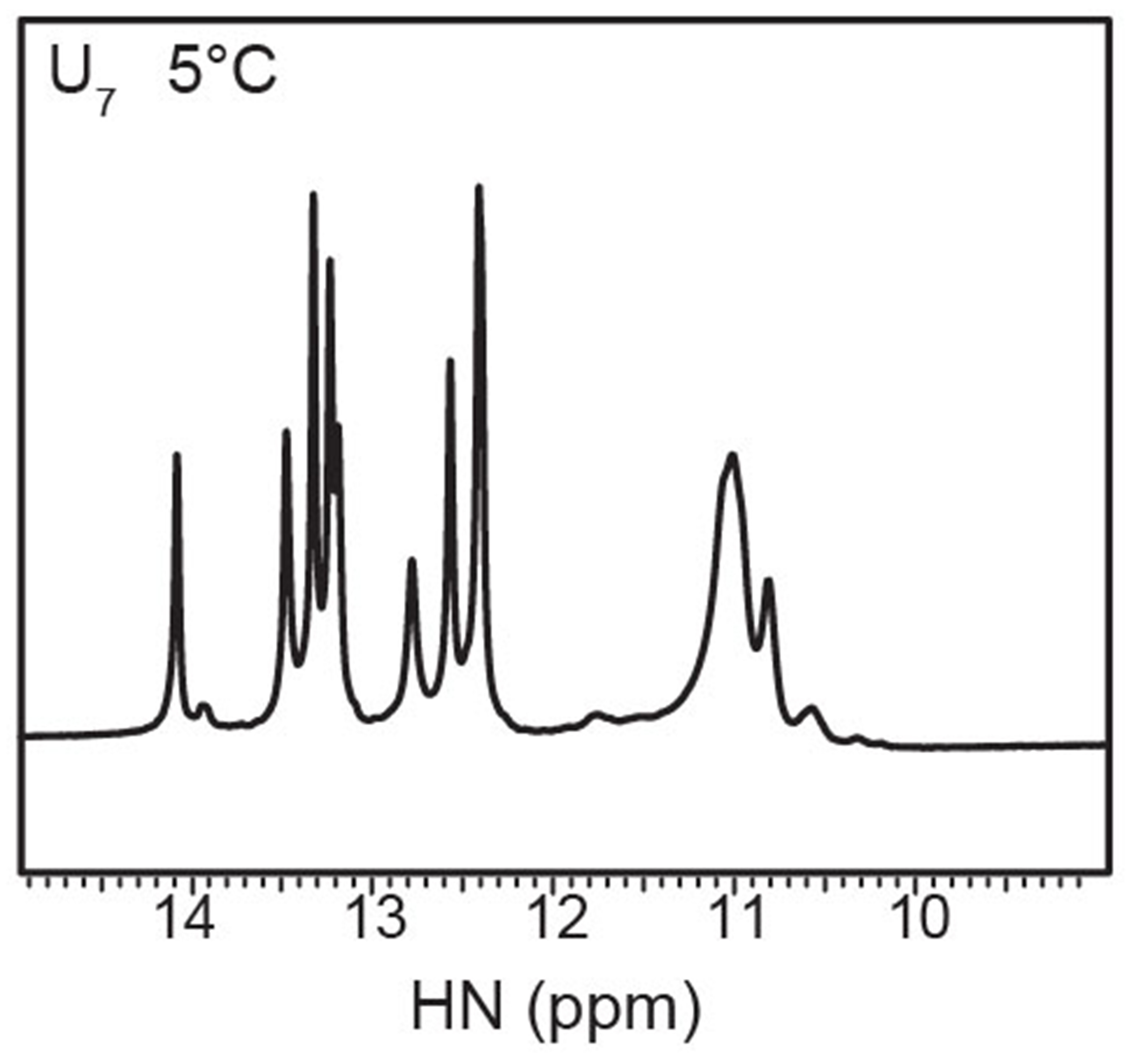
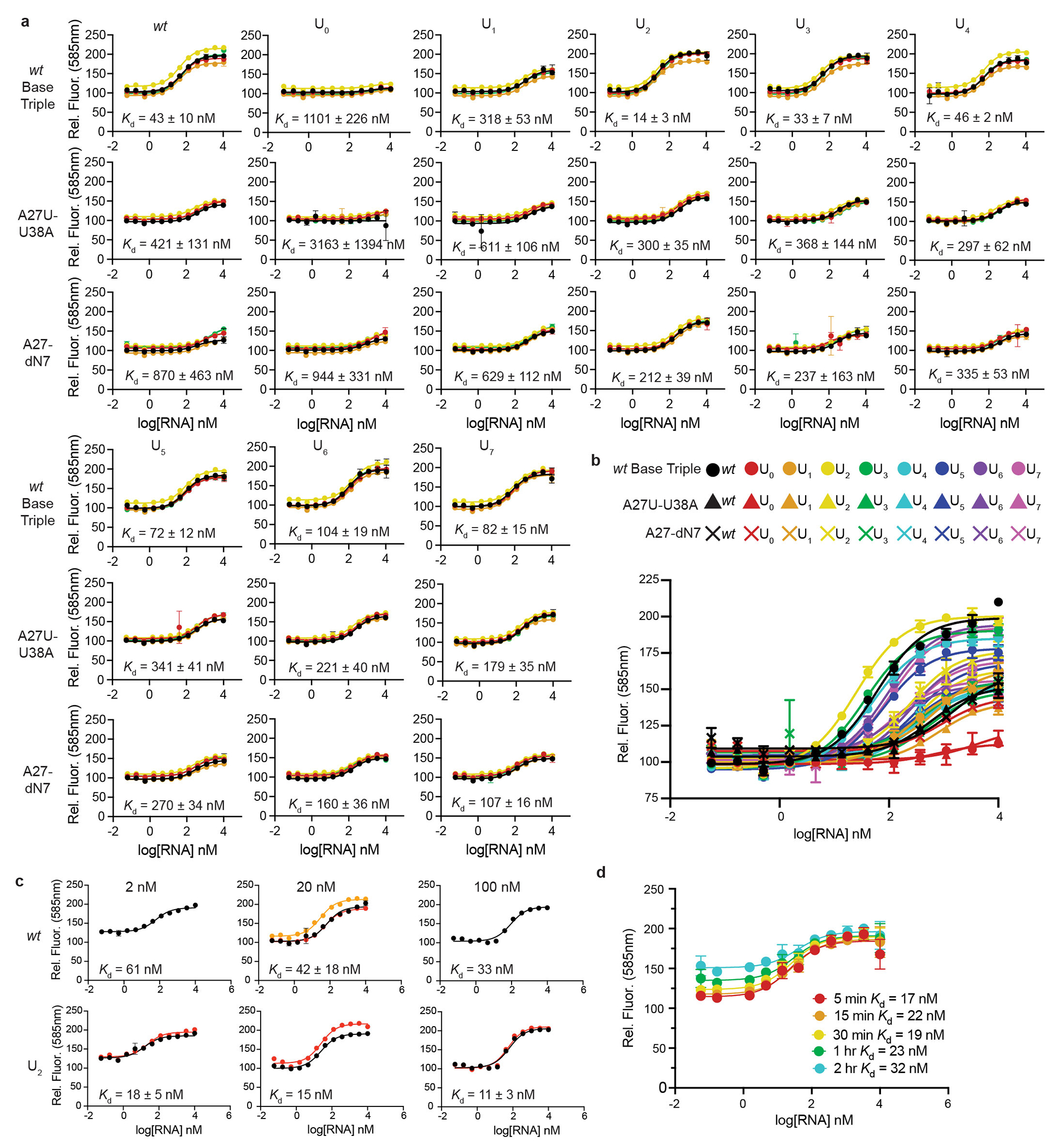
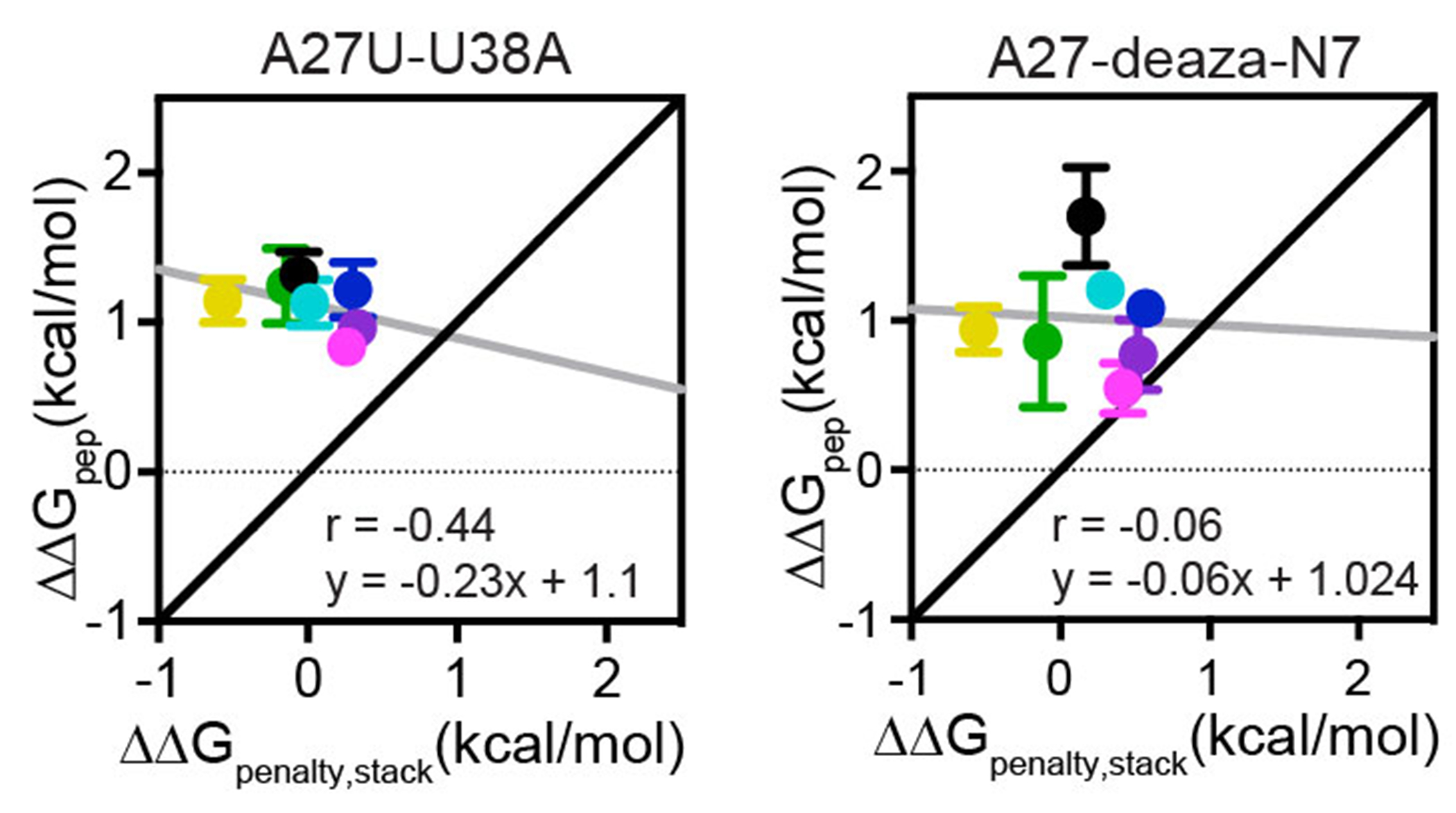
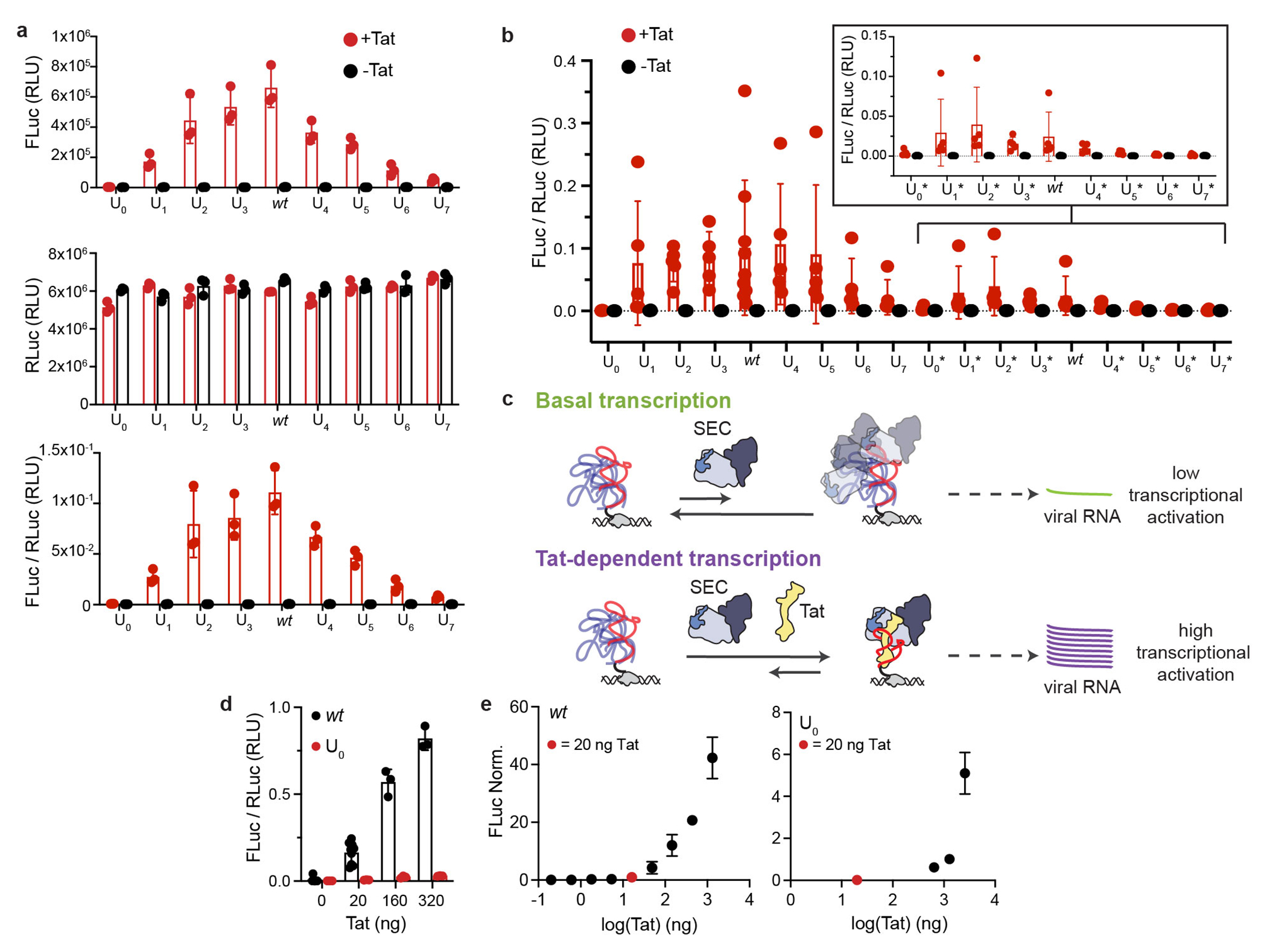

References
-
- Parvin JD, Mc Cormick RJ, Sharp PA & Fisher DE Pre-bending of a promoter sequence enhances affinity for the TATA-binding factor. Nature 373, 727 (1995). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
