

SARS-CoV-2 Nucleocapsid Protein Is a Potential Therapeutic Target for Anticoronavirus Drug Discovery
- PMID: 37199631
- PMCID: PMC10269701
- DOI: 10.1128/spectrum.01186-23
SARS-CoV-2 Nucleocapsid Protein Is a Potential Therapeutic Target for Anticoronavirus Drug Discovery
Abstract
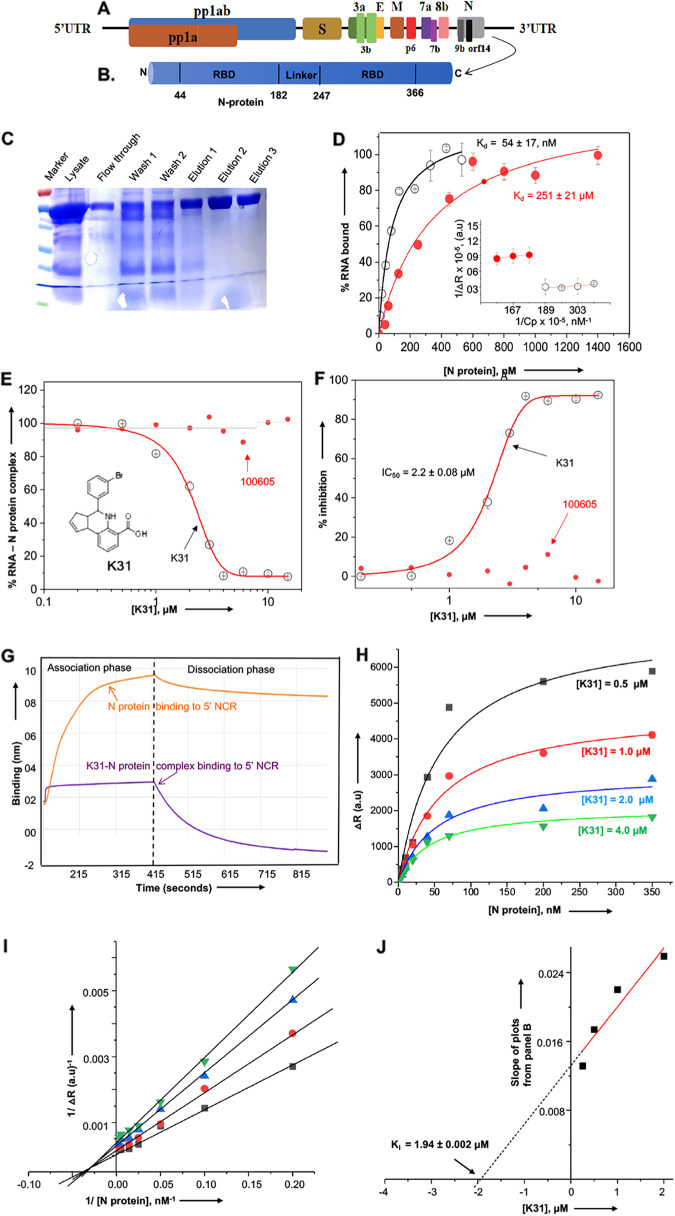
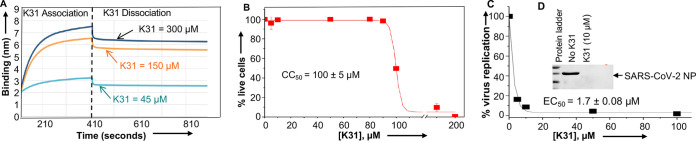
SARS-CoV-2, the etiologic agent of the COVID-19 pandemic, is a highly contagious positive-sense RNA virus. Its explosive community spread and the emergence of new mutant strains have created palpable anxiety even in vaccinated people. The lack of effective anticoronavirus therapeutics continues to be a major global health concern, especially due to the high evolution rate of SARS-CoV-2. The nucleocapsid protein (N protein) of SARS-CoV-2 is highly conserved and involved in diverse processes of the virus replication cycle. Despite its critical role in coronavirus replication, N protein remains an unexplored target for anticoronavirus drug discovery. Here, we demonstrate that a novel compound, K31, binds to the N protein of SARS-CoV-2 and noncompetitively inhibits its binding to the 5' terminus of the viral genomic RNA. K31 is well tolerated by SARS-CoV-2-permissive Caco2 cells. Our results show that K31 inhibited SARS-CoV-2 replication in Caco2 cells with a selective index of ~58. These observations suggest that SARS-CoV-2 N protein is a druggable target for anticoronavirus drug discovery. K31 holds promise for further development as an anticoronavirus therapeutic. IMPORTANCE The lack of potent antiviral drugs for SARS-CoV-2 is a serious global health concern, especially with the explosive spread of the COVID-19 pandemic worldwide and the constant emergence of new mutant strains with improved human-to-human transmission. Although an effective coronavirus vaccine appears promising, the lengthy vaccine development processes in general and the emergence of new mutant viral strains with a potential to evade the vaccine always remain a serious concern. The antiviral drugs targeted to the highly conserved targets of viral or host origin remain the most viable and timely approach, easily accessible to the general population, in combating any new viral illness. The majority of anticoronavirus drug development efforts have focused on spike protein, envelope protein, 3CLpro, and Mpro. Our results show that virus-encoded N protein is a novel therapeutic target for anticoronavirus drug discovery. Due to its high conservation, the anti-N protein inhibitors will likely have broad-spectrum anticoronavirus activity.
Keywords: RNA virus; antiviral agents; coronavirus; nucleocapsid protein; virus replication.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Wu A, Peng Y, Huang B, Ding X, Wang X, Niu P, Meng J, Zhu Z, Zhang Z, Wang J, Sheng J, Quan L, Xia Z, Tan W, Cheng G, Jiang T. 2020. Genome composition and divergence of the novel coronavirus (2019-nCoV) originating in China. Cell Host Microbe 27:325–328. doi:10.1016/j.chom.2020.02.001. - DOI - PMC - PubMed
-
- Siu YL, Teoh KT, Lo J, Chan CM, Kien F, Escriou N, Tsao SW, Nicholls JM, Altmeyer R, Peiris JS, Bruzzone R, Nal B. 2008. The M, E, and N structural proteins of the severe acute respiratory syndrome coronavirus are required for efficient assembly, trafficking, and release of virus-like particles. J Virol 82:11318–11330. doi:10.1128/JVI.01052-08. - DOI - PMC - PubMed
-
- Surjit M, Kumar R, Mishra RN, Reddy MK, Chow VT, Lal SK. 2005. The severe acute respiratory syndrome coronavirus nucleocapsid protein is phosphorylated and localizes in the cytoplasm by 14-3-3-mediated translocation. J Virol 79:11476–11486. doi:10.1128/JVI.79.17.11476-11486.2005. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
