

Conserved stromal-immune cell circuits secure B cell homeostasis and function
- PMID: 37202489
- PMCID: PMC10307622
- DOI: 10.1038/s41590-023-01503-3
Conserved stromal-immune cell circuits secure B cell homeostasis and function
Abstract
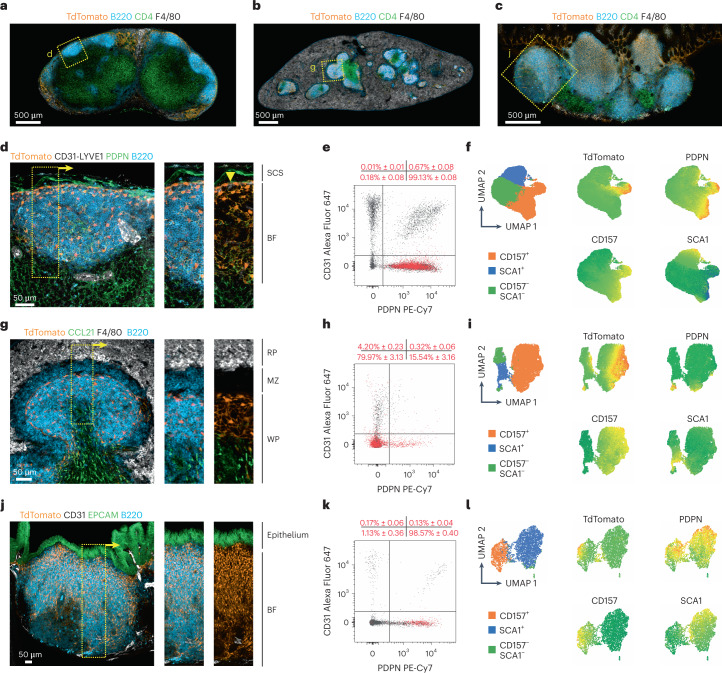
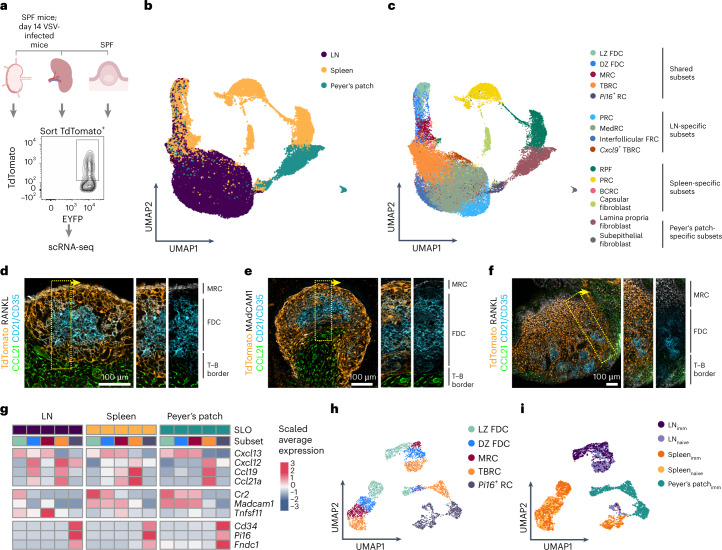
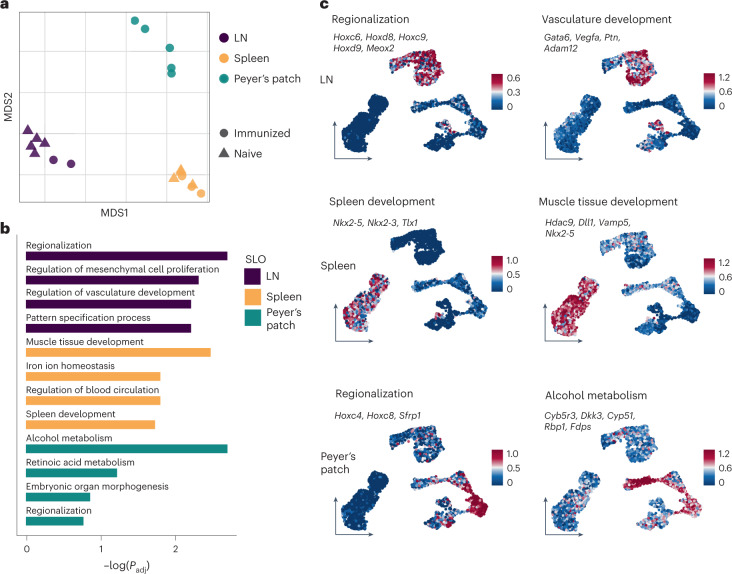
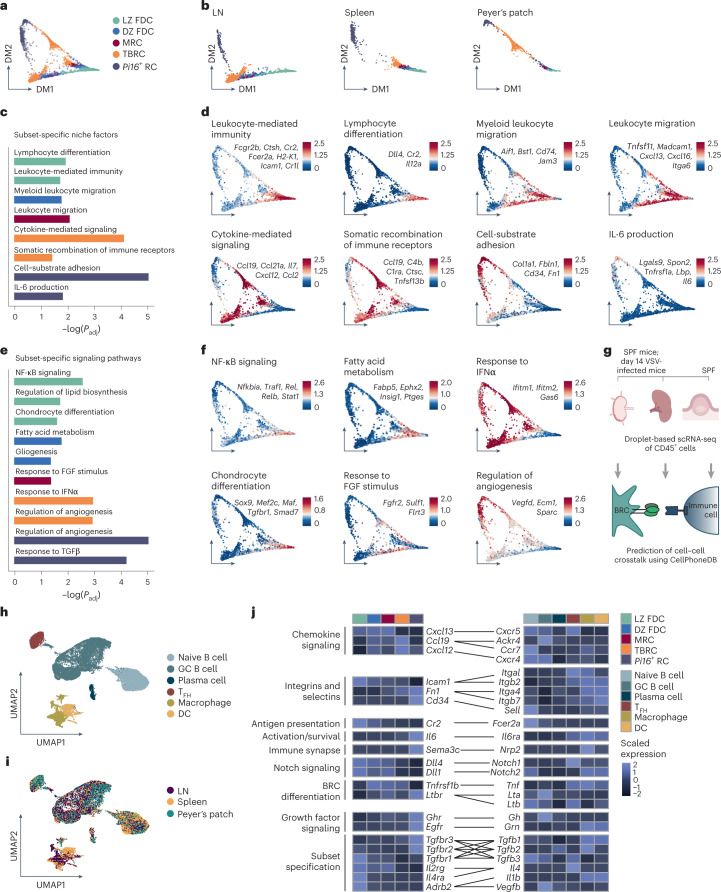
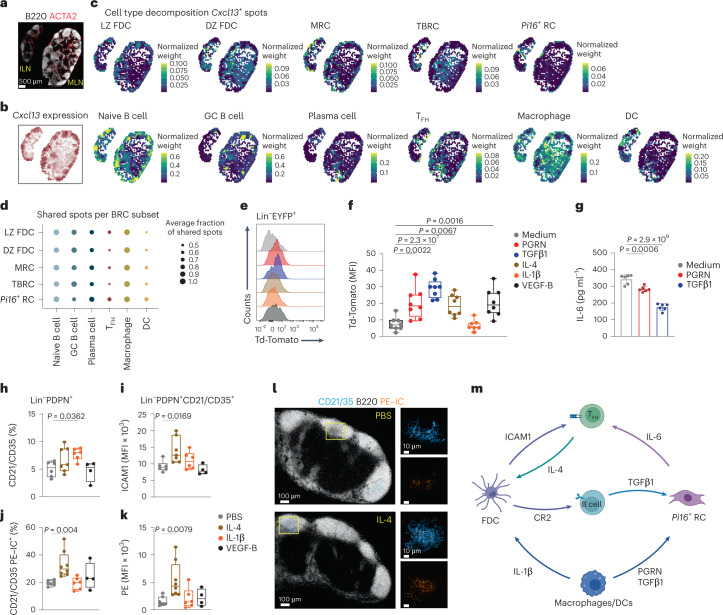
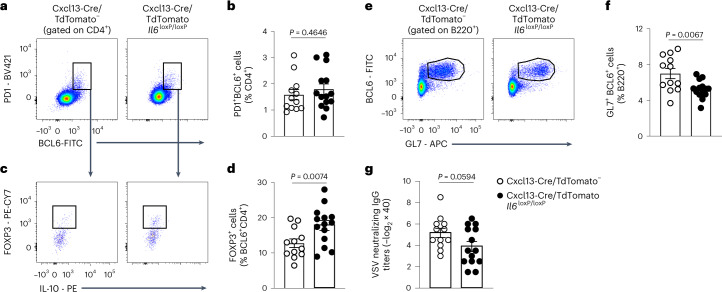
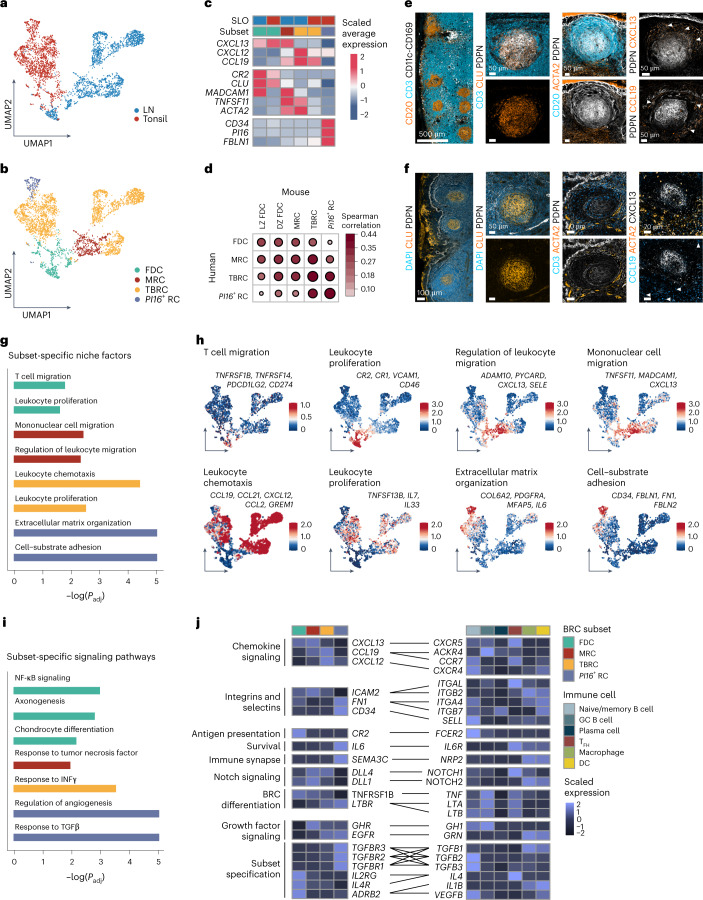
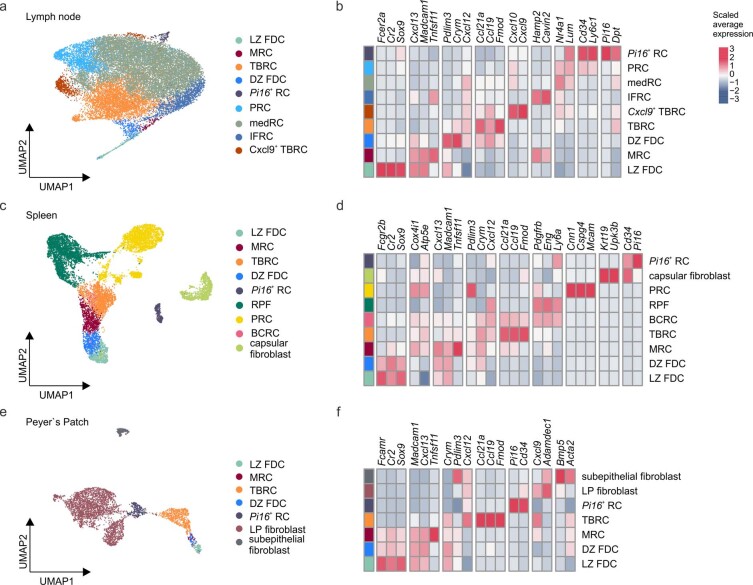
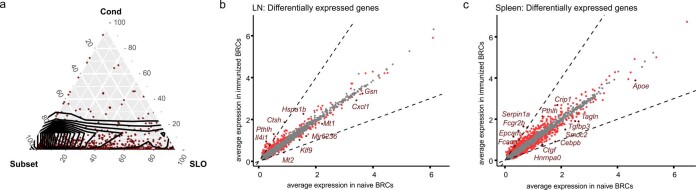
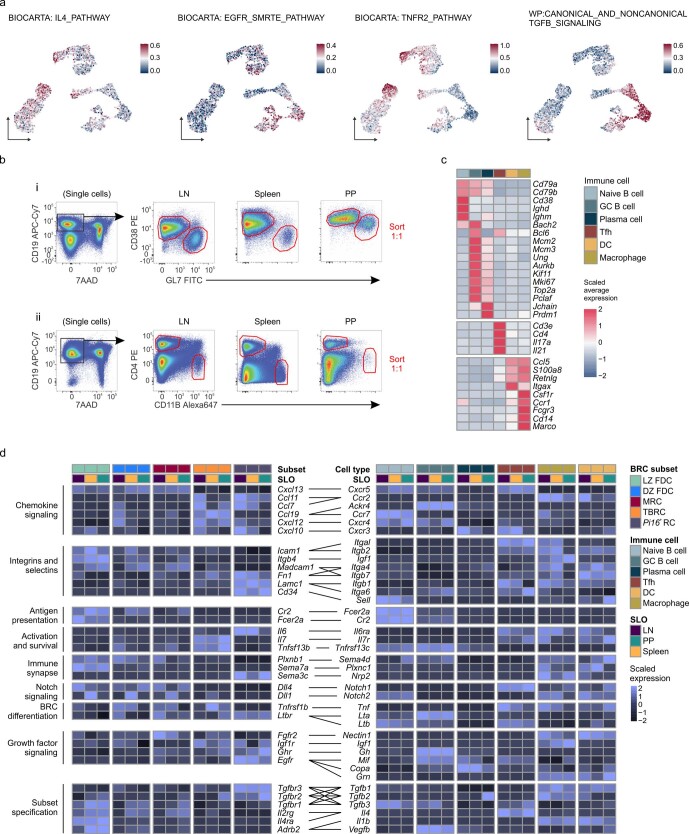
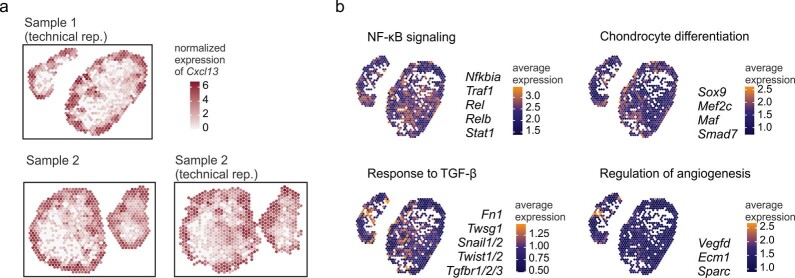
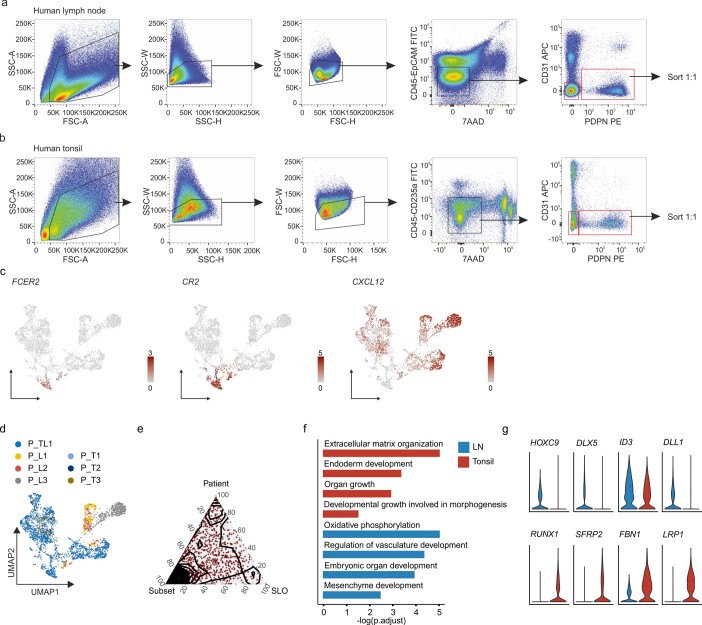
B cell zone reticular cells (BRCs) form stable microenvironments that direct efficient humoral immunity with B cell priming and memory maintenance being orchestrated across lymphoid organs. However, a comprehensive understanding of systemic humoral immunity is hampered by the lack of knowledge of global BRC sustenance, function and major pathways controlling BRC-immune cell interactions. Here we dissected the BRC landscape and immune cell interactome in human and murine lymphoid organs. In addition to the major BRC subsets underpinning the follicle, including follicular dendritic cells, PI16+ RCs were present across organs and species. As well as BRC-produced niche factors, immune cell-driven BRC differentiation and activation programs governed the convergence of shared BRC subsets, overwriting tissue-specific gene signatures. Our data reveal that a canonical set of immune cell-provided cues enforce bidirectional signaling programs that sustain functional BRC niches across lymphoid organs and species, thereby securing efficient humoral immunity.
© 2023. The Author(s).
Conflict of interest statement
C.P.-S., C.G.-C., H.-W.C., L.O., N.B.P. and B.L. are founders and shareholders of Stromal Therapeutics. L.O. and B.L. are members of the board of Stromal Therapeutics. The other authors declare no competing interests.
Figures




Comment in
-
New fibroblast network connections support lymphocytic cellular service.Nat Immunol. 2023 Jul;24(7):1054-1055. doi: 10.1038/s41590-023-01537-7. Nat Immunol. 2023. PMID: 37322180 No abstract available.
References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
