

Interneuronal GluK1 kainate receptors control maturation of GABAergic transmission and network synchrony in the hippocampus
- PMID: 37210550
- PMCID: PMC10199616
- DOI: 10.1186/s13041-023-01035-9
Interneuronal GluK1 kainate receptors control maturation of GABAergic transmission and network synchrony in the hippocampus
Abstract
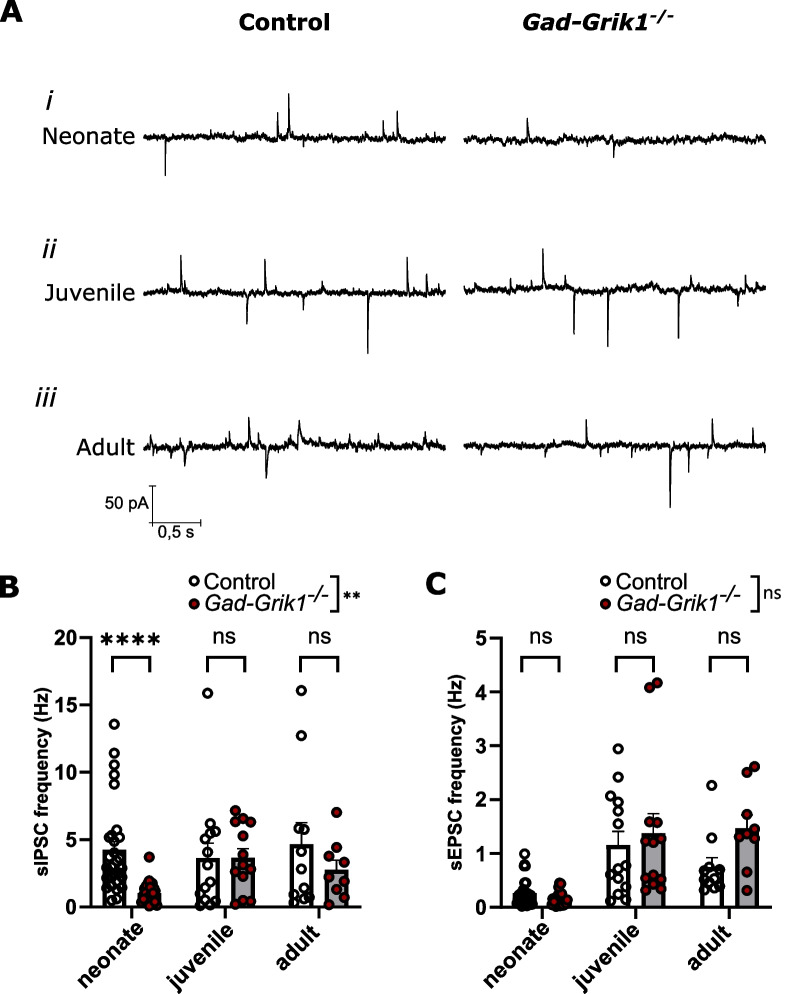
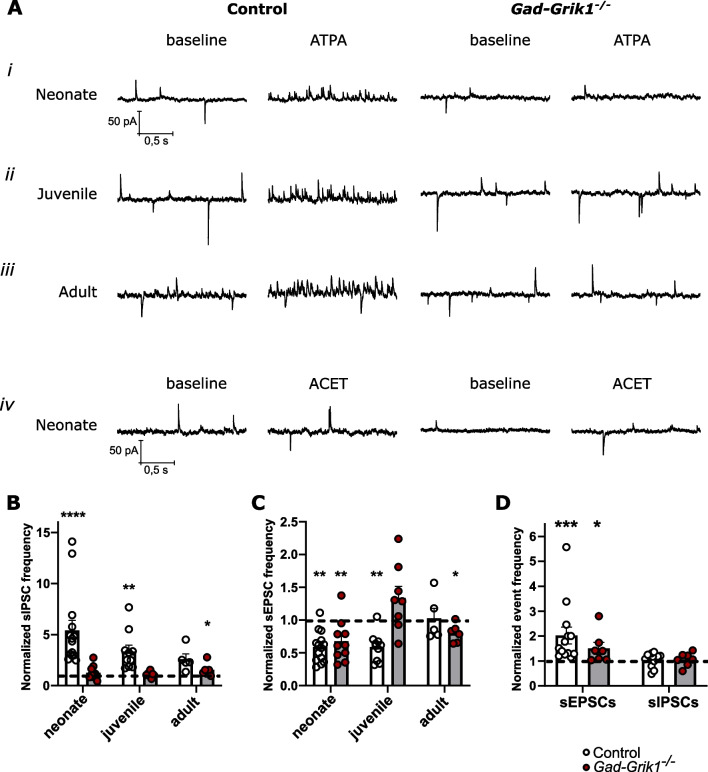
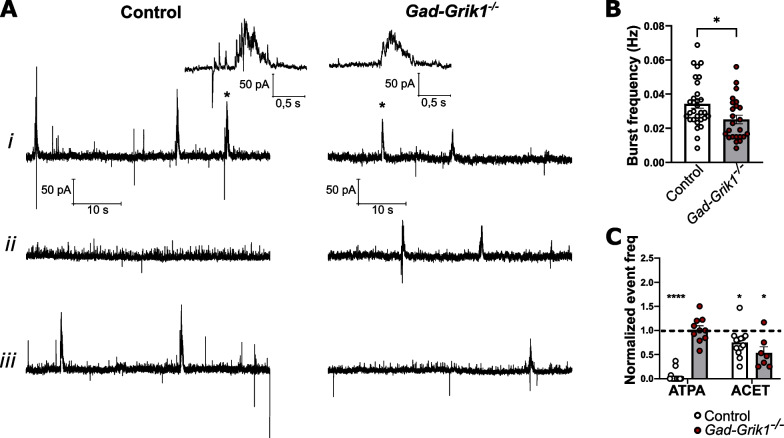
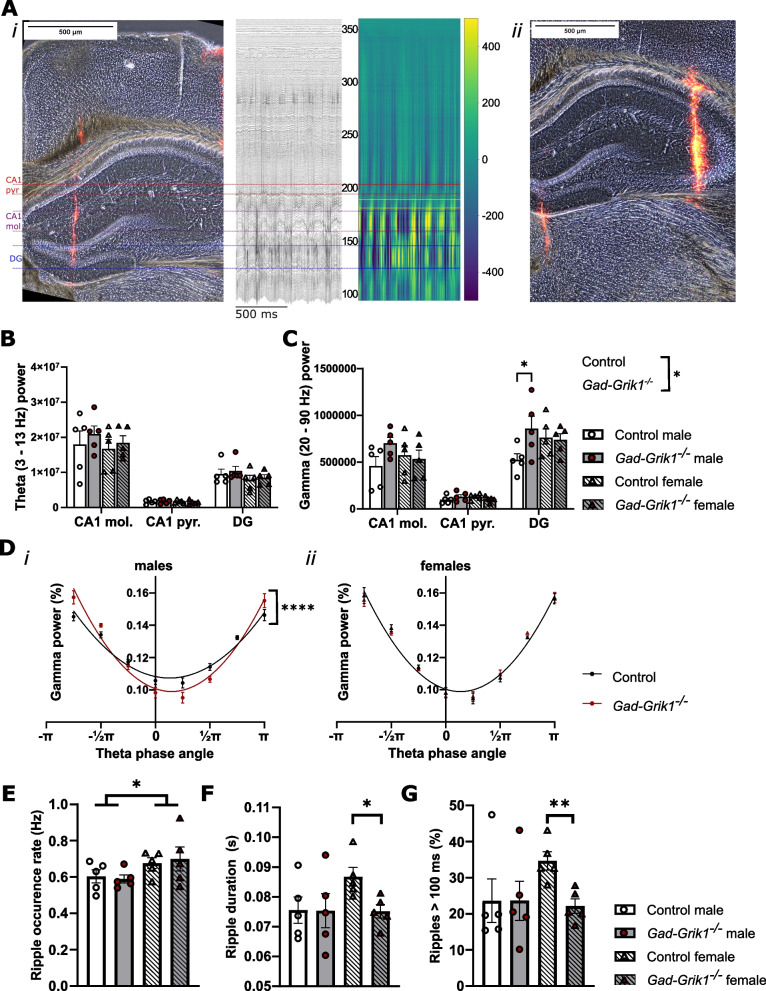
Kainate type glutamate receptors (KARs) are strongly expressed in GABAergic interneurons and have the capability of modulating their functions via ionotropic and G-protein coupled mechanisms. GABAergic interneurons are critical for generation of coordinated network activity in both neonatal and adult brain, yet the role of interneuronal KARs in network synchronization remains unclear. Here, we show that GABAergic neurotransmission and spontaneous network activity is perturbed in the hippocampus of neonatal mice lacking GluK1 KARs selectively in GABAergic neurons. Endogenous activity of interneuronal GluK1 KARs maintains the frequency and duration of spontaneous neonatal network bursts and restrains their propagation through the hippocampal network. In adult male mice, the absence of GluK1 in GABAergic neurons led to stronger hippocampal gamma oscillations and enhanced theta-gamma cross frequency coupling, coinciding with faster spatial relearning in the Barnes maze. In females, loss of interneuronal GluK1 resulted in shorter sharp wave ripple oscillations and slightly impaired abilities in flexible sequencing task. In addition, ablation of interneuronal GluK1 resulted in lower general activity and novel object avoidance, while causing only minor anxiety phenotype. These data indicate a critical role for GluK1 containing KARs in GABAergic interneurons in regulation of physiological network dynamics in the hippocampus at different stages of development.
Keywords: Cognitive flexibility; GABAergic interneuron; Gamma oscillation; Glutamate receptor; Hippocampus; Kainate receptor; Network synchronization.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures

References
-
- Duan ZRS, Che A, Chu P, Modol L, Bollmann Y, Babij R, Fetcho RN, Otsuka T, Fuccillo MV, Liston C, Pisapia DJ, Cossart R, De Marco García NV. GABAergic restriction of network dynamics regulates interneuron survival in the developing cortex. Neuron. 2020;105(1):75–92.e75. doi: 10.1016/j.neuron.2019.10.008. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
