

Cold atmospheric pressure plasma-antibiotic synergy in Pseudomonas aeruginosa biofilms is mediated via oxidative stress response
- PMID: 37214348
- PMCID: PMC10196807
- DOI: 10.1016/j.bioflm.2023.100122
Cold atmospheric pressure plasma-antibiotic synergy in Pseudomonas aeruginosa biofilms is mediated via oxidative stress response
Abstract
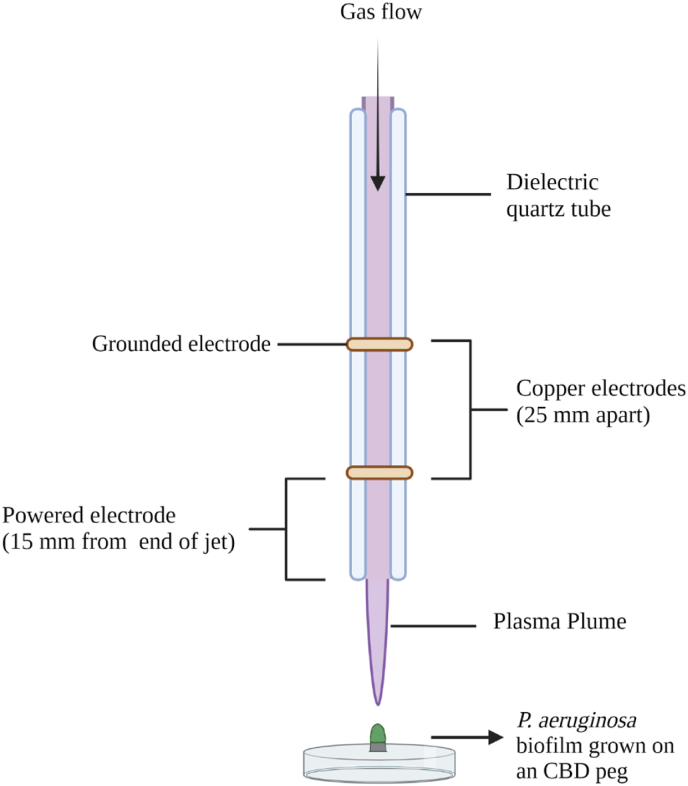
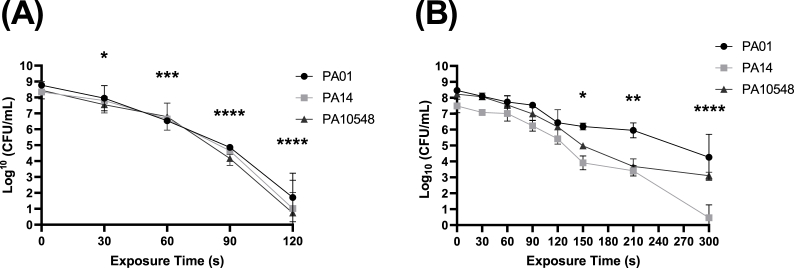
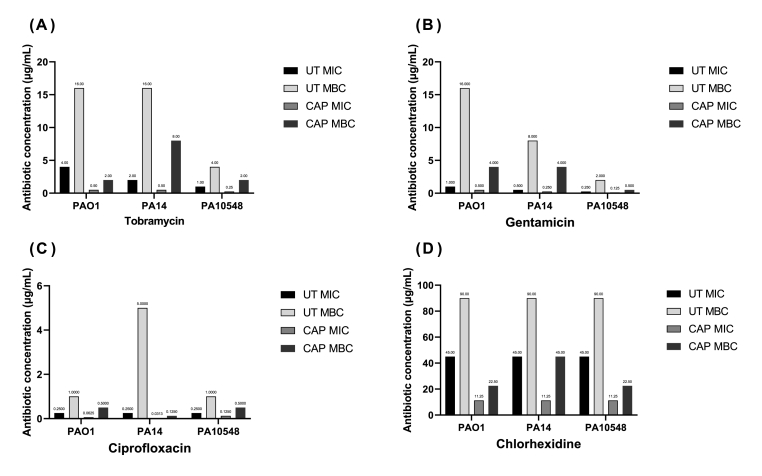
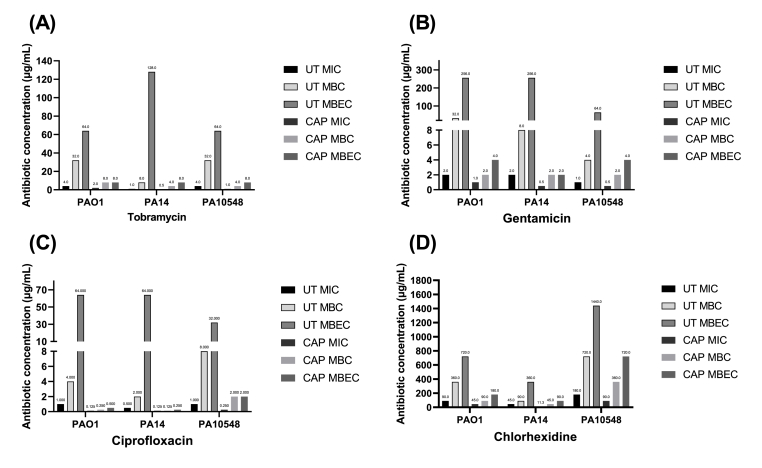
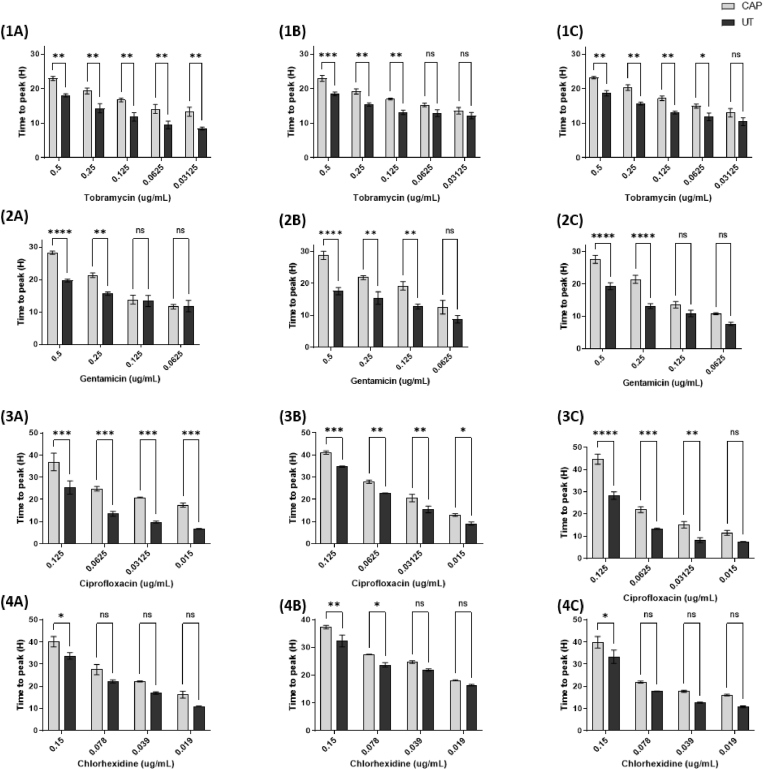
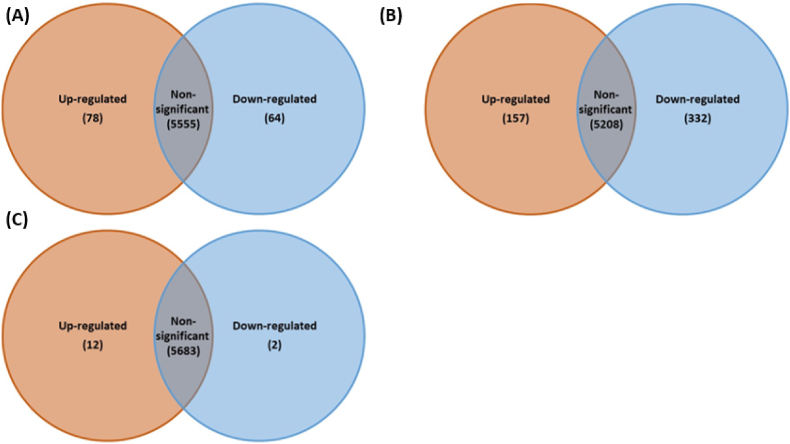
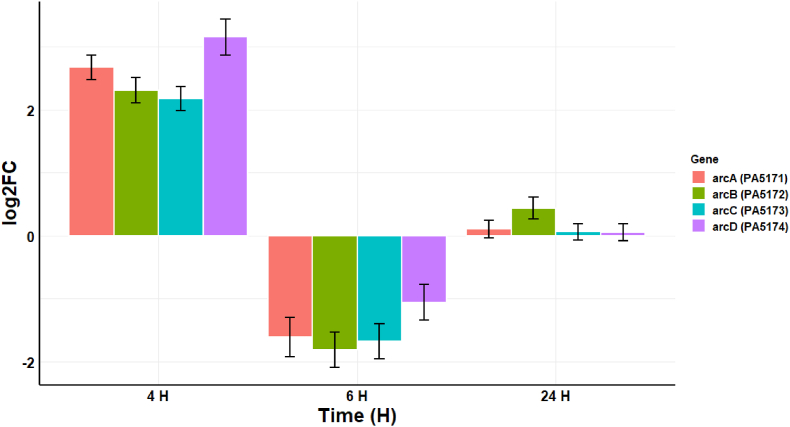
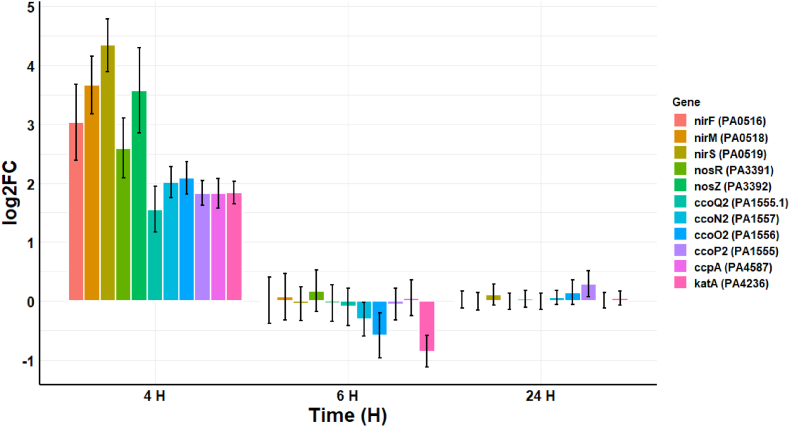
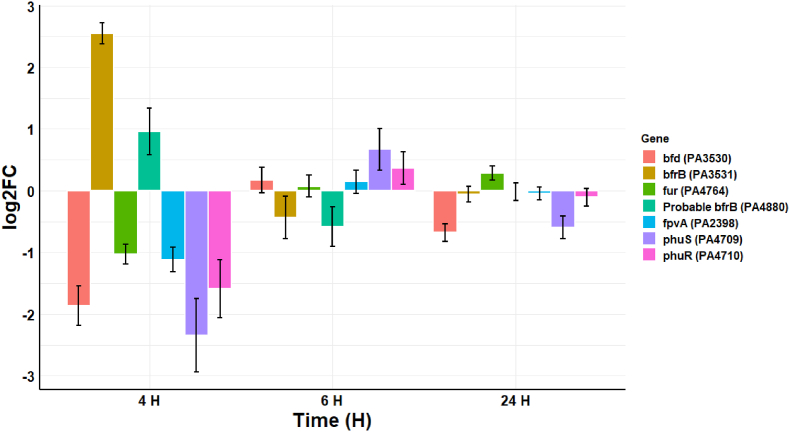
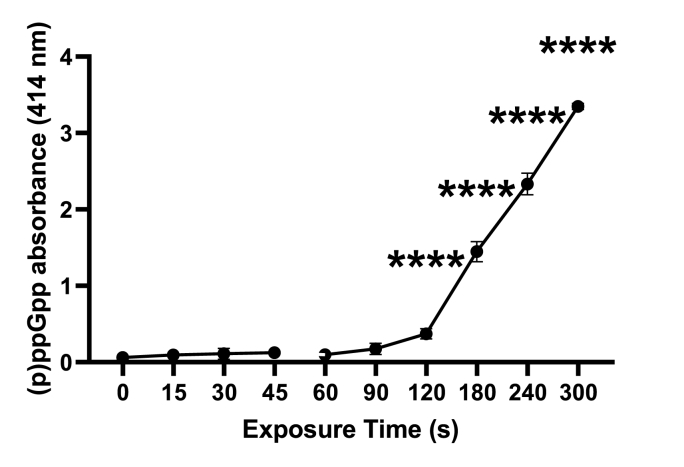
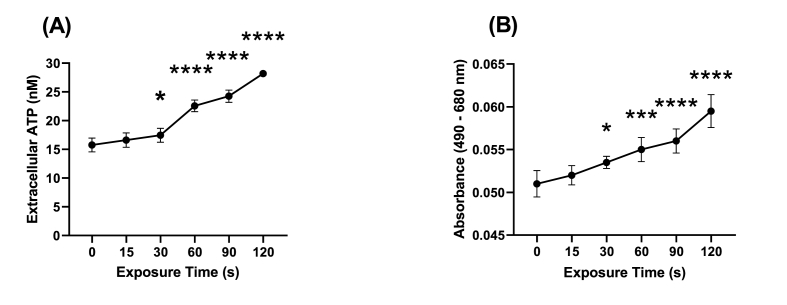
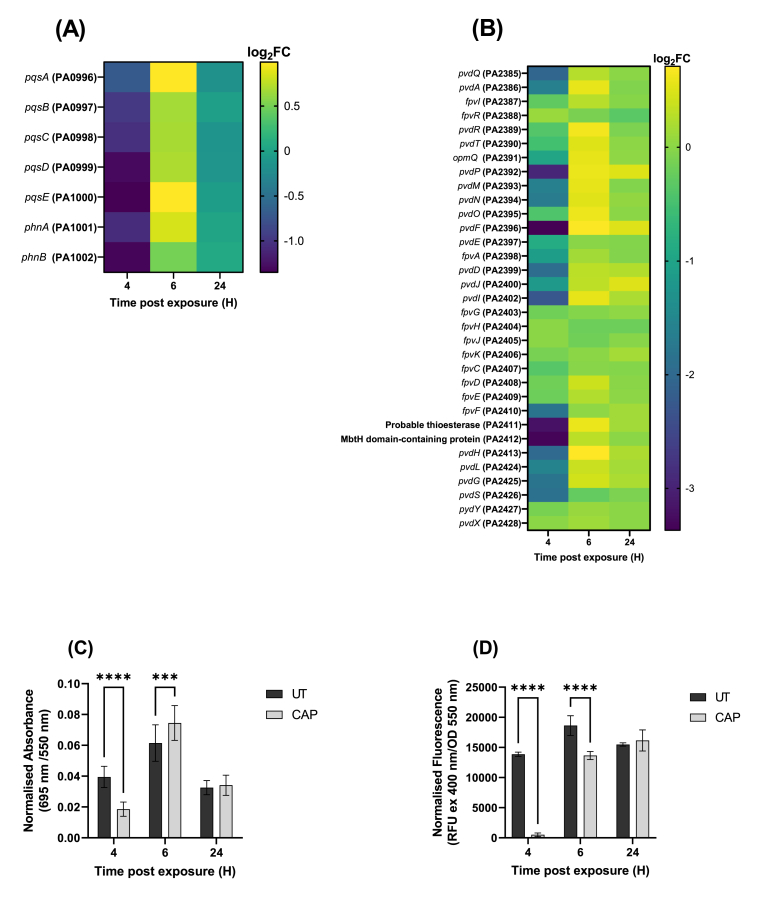
Cold atmospheric-pressure plasma (CAP) has emerged as a potential alternative or adjuvant to conventional antibiotics for the treatment of bacterial infections, including those caused by antibiotic-resistant pathogens. The potential of sub-lethal CAP exposures to synergise conventional antimicrobials for the eradication of Pseudomonas aeruginosa biofilms is investigated in this study. The efficacy of antimicrobials following or in the absence of sub-lethal CAP pre-treatment in P. aeruginosa biofilms was assessed. CAP pre-treatment resulted in an increase in both planktonic and biofilm antimicrobial sensitivity for all three strains tested (PAO1, PA14, and PA10548), with both minimum inhibitory concentrations (MICs) and minimum biofilm eradication concentrations (MBECs) of individual antimicrobials, being significantly reduced following CAP pre-treatment of the biofilm (512-fold reduction with ciprofloxacin/gentamicin; and a 256-fold reduction with tobramycin). At all concentrations of antimicrobial used, the combination of sub-lethal CAP exposure and antimicrobials was effective at increasing time-to-peak metabolism, as measured by isothermal microcalorimetry, again indicating enhanced susceptibility. CAP is known to damage bacterial cell membranes and DNA by causing oxidative stress through the in situ generation of reactive oxygen and nitrogen species (RONS). While the exact mechanism is not clear, oxidative stress on outer membrane proteins is thought to damage/perturb cell membranes, confirmed by ATP and LDH leakage, allowing antimicrobials to penetrate the bacterial cell more effectively, thus increasing bacterial susceptibility. Transcriptomic analysis, reveals that cold-plasma mediated oxidative stress caused upregulation of P. aeruginosa superoxide dismutase, cbb3 oxidases, catalases, and peroxidases, and upregulation in denitrification genes, suggesting that P. aeruginosa uses these enzymes to degrade RONS and mitigate the effects of cold plasma mediated oxidative stress. CAP treatment also led to an increased production of the signalling molecule ppGpp in P. aeruginosa, indicative of a stringent response being established. Although we did not directly measure persister cell formation, this stringent response may potentially be associated with the formation of persister cells in biofilm cultures. The production of ppGpp and polyphosphate may be associated with protein synthesis inhibition and increase efflux pump activity, factors which can result in antimicrobial tolerance. The transcriptomic analysis also showed that by 6 h post-treatment, there was downregulation in ribosome modulation factor, which is involved in the formation of persister cells, suggesting that the cells had begun to resuscitate/recover. In addition, CAP treatment at 4 h post-exposure caused downregulation of the virulence factors pyoverdine and pyocyanin; by 6 h post-exposure, virulence factor production was increasing. Transcriptomic analysis provides valuable insights into the mechanisms by which P. aeruginosa biofilms exhibits enhanced susceptibility to antimicrobials. Overall, these findings suggest, for the first time, that short CAP sub-lethal pre-treatment can be an effective strategy for enhancing the susceptibility of P. aeruginosa biofilms to antimicrobials and provides important mechanistic insights into cold plasma-antimicrobial synergy. Transcriptomic analysis of the response to, and recovery from, sub-lethal cold plasma exposures in P. aeruginosa biofilms improves our current understanding of cold plasma biofilm interactions.
Keywords: Antimicrobial synergy; Biofilm; Cold plasma; Persister cells; Plasma Medicine; Pseudomonas aeruginosa; Transcriptomics; ppGpp.
© 2023 Published by Elsevier B.V.
Conflict of interest statement
The authors of this manuscript declare that they have no financial or personal relationships that may be perceived as influencing their work. We confirm that all authors have contributed significantly to the planning, execution, and writing of the manuscript. We also confirm that the content of this manuscript has not been published or submitted for publication elsewhere.
Figures
References
-
- Flynn P.B., Higginbotham S., Nid’a H.A., Gorman S.P., Graham W.G., Gilmore B.F. Bactericidal efficacy of atmospheric pressure non-thermal plasma (APNTP) against the ESKAPE pathogens. Int J Antimicrob Agents. 2015;46(1):101–107. - PubMed
-
- Lewis K. Persister cells. Annu Rev Microbiol. 2010;64:357–372. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
