

This is a preprint.
Pyruvate dehydrogenase complex integrates the metabolome and epigenome in CD8+ memory T cell differentiation in vitro
- PMID: 37215014
- PMCID: PMC10197744
- DOI: 10.21203/rs.3.rs-2838359/v1
Pyruvate dehydrogenase complex integrates the metabolome and epigenome in CD8+ memory T cell differentiation in vitro
Abstract
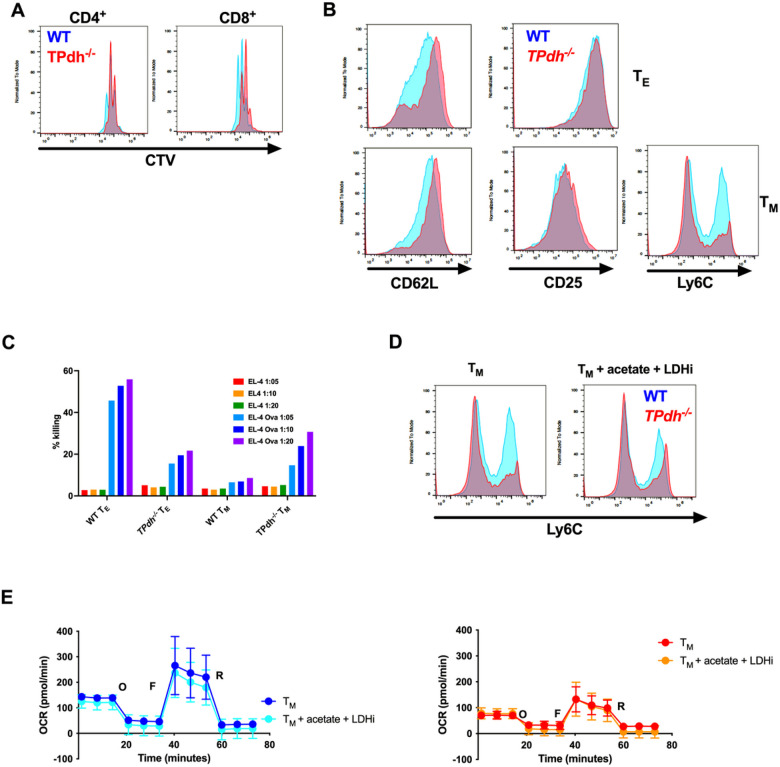
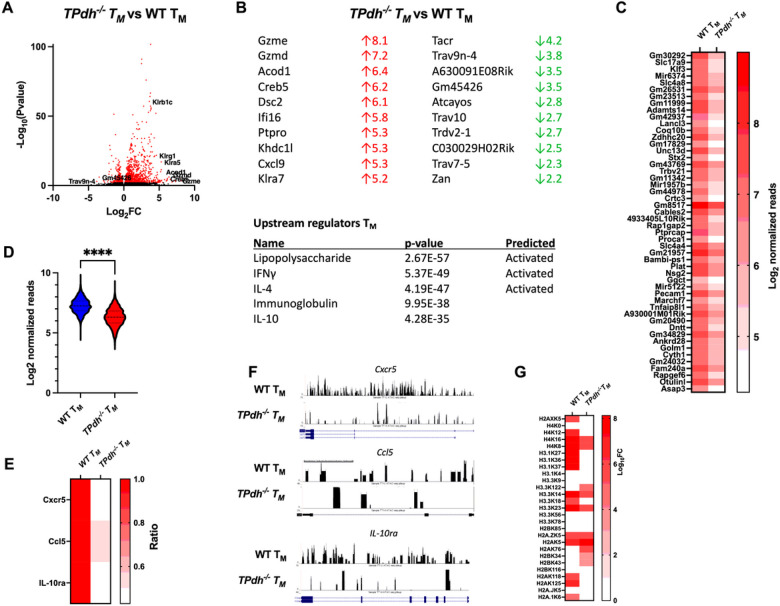
Modulation of metabolic flux through pyruvate dehydrogenase complex (PDC) plays an important role in T cell activation and differentiation. PDC sits at the transition between glycolysis and the tricarboxylic acid cycle and is a major producer of acetyl-CoA, marking it as a potential metabolic and epigenetic node To understand the role of pyruvate dehydrogenase complex in T cell differentiation, we generated mice deficient in T cell pyruvate dehydrogenase E1A (Pdha) subunit using a CD4-cre recombinase-based strategy. Herein, we show that genetic ablation of PDC activity in T cells (TPdh-/-) leads to marked perturbations in glycolysis, the tricarboxylic acid cycle, and OXPHOS. TPdh-/- T cells became dependent upon substrate level phosphorylation via glycolysis, secondary to depressed OXPHOS. Due to the block of PDC activity, histone acetylation was also reduced, including H3K27, a critical site for CD8+ TM differentiation. Transcriptional and functional profiling revealed abnormal CD8+ TM differentiation in vitro. Collectively, our data indicate that PDC integrates the metabolome and epigenome in CD8+ memory T cell differentiation. Targeting this metabolic and epigenetic node can have widespread ramifications on cellular function.
Figures



References
Publication types
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
