

Cortical interneurons: fit for function and fit to function? Evidence from development and evolution
- PMID: 37215503
- PMCID: PMC10192557
- DOI: 10.3389/fncir.2023.1172464
Cortical interneurons: fit for function and fit to function? Evidence from development and evolution
Abstract
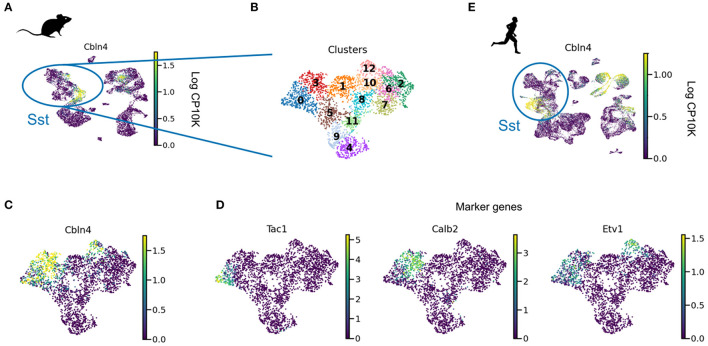
Cortical inhibitory interneurons form a broad spectrum of subtypes. This diversity suggests a division of labor, in which each cell type supports a distinct function. In the present era of optimisation-based algorithms, it is tempting to speculate that these functions were the evolutionary or developmental driving force for the spectrum of interneurons we see in the mature mammalian brain. In this study, we evaluated this hypothesis using the two most common interneuron types, parvalbumin (PV) and somatostatin (SST) expressing cells, as examples. PV and SST interneurons control the activity in the cell bodies and the apical dendrites of excitatory pyramidal cells, respectively, due to a combination of anatomical and synaptic properties. But was this compartment-specific inhibition indeed the function for which PV and SST cells originally evolved? Does the compartmental structure of pyramidal cells shape the diversification of PV and SST interneurons over development? To address these questions, we reviewed and reanalyzed publicly available data on the development and evolution of PV and SST interneurons on one hand, and pyramidal cell morphology on the other. These data speak against the idea that the compartment structure of pyramidal cells drove the diversification into PV and SST interneurons. In particular, pyramidal cells mature late, while interneurons are likely committed to a particular fate (PV vs. SST) during early development. Moreover, comparative anatomy and single cell RNA-sequencing data indicate that PV and SST cells, but not the compartment structure of pyramidal cells, existed in the last common ancestor of mammals and reptiles. Specifically, turtle and songbird SST cells also express the Elfn1 and Cbln4 genes that are thought to play a role in compartment-specific inhibition in mammals. PV and SST cells therefore evolved and developed the properties that allow them to provide compartment-specific inhibition before there was selective pressure for this function. This suggest that interneuron diversity originally resulted from a different evolutionary driving force and was only later co-opted for the compartment-specific inhibition it seems to serve in mammals today. Future experiments could further test this idea using our computational reconstruction of ancestral Elfn1 protein sequences.
Keywords: development; evolution; inhibition; interneuron; microcircuits; neural morphology; pyramidal cell dendrites; single cell RNA seq.
Copyright © 2023 Keijser and Sprekeler.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures








Similar articles
-
Inhibition by Somatostatin Interneurons in Olfactory Cortex.Front Neural Circuits. 2016 Aug 17;10:62. doi: 10.3389/fncir.2016.00062. eCollection 2016. Front Neural Circuits. 2016. PMID: 27582691 Free PMC article.
-
Parvalbumin and Somatostatin Interneurons Contribute to the Generation of Hippocampal Gamma Oscillations.J Neurosci. 2020 Sep 30;40(40):7668-7687. doi: 10.1523/JNEUROSCI.0261-20.2020. Epub 2020 Aug 28. J Neurosci. 2020. PMID: 32859716 Free PMC article.
-
The Impact of SST and PV Interneurons on Nonlinear Synaptic Integration in the Neocortex.eNeuro. 2021 Sep 7;8(5):ENEURO.0235-21.2021. doi: 10.1523/ENEURO.0235-21.2021. Print 2021 Sep-Oct. eNeuro. 2021. PMID: 34400470 Free PMC article.
-
Reliable Sensory Processing in Mouse Visual Cortex through Cooperative Interactions between Somatostatin and Parvalbumin Interneurons.J Neurosci. 2021 Oct 20;41(42):8761-8778. doi: 10.1523/JNEUROSCI.3176-20.2021. Epub 2021 Sep 7. J Neurosci. 2021. PMID: 34493543 Free PMC article.
-
Distinct FGF-FGFR sets regulate inhibitory presynaptic differentiation from parvalbumin- and somatostatin-positive interneurons.Development. 2025 Jul 1;152(13):dev204532. doi: 10.1242/dev.204532. Epub 2025 Jul 3. Development. 2025. PMID: 40554746
Cited by
-
Perturbing cortical networks: in vivo electrophysiological consequences of pan-neuronal chemogenetic manipulations using deschloroclozapine.Front Neurosci. 2024 Apr 25;18:1396978. doi: 10.3389/fnins.2024.1396978. eCollection 2024. Front Neurosci. 2024. PMID: 38726028 Free PMC article.
-
The Radioprotective Effect of LBP on Neurogenesis and Cognition after Acute Radiation Exposure.Curr Radiopharm. 2024;17(3):257-265. doi: 10.2174/0118744710274008231220055033. Curr Radiopharm. 2024. PMID: 38204264 Free PMC article.
-
Transcriptomic Correlates of State Modulation in GABAergic Interneurons: A Cross-Species Analysis.J Neurosci. 2024 Oct 30;44(44):e2371232024. doi: 10.1523/JNEUROSCI.2371-23.2024. J Neurosci. 2024. PMID: 39299800 Free PMC article.
-
Novel pharmacological targets for GABAergic dysfunction in ADHD.Neuropharmacology. 2024 May 15;249:109897. doi: 10.1016/j.neuropharm.2024.109897. Epub 2024 Mar 8. Neuropharmacology. 2024. PMID: 38462041 Free PMC article. Review.
-
Interleaving asynchronous and synchronous activity in balanced cortical networks with short-term synaptic depression.bioRxiv [Preprint]. 2025 Mar 18:2025.03.13.643074. doi: 10.1101/2025.03.13.643074. bioRxiv. 2025. PMID: 40166256 Free PMC article. Preprint.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources