

Intrinsic suppression of type I interferon production underlies the therapeutic efficacy of IL-15-producing natural killer cells in B-cell acute lymphoblastic leukemia
- PMID: 37217248
- PMCID: PMC10231005
- DOI: 10.1136/jitc-2022-006649
Intrinsic suppression of type I interferon production underlies the therapeutic efficacy of IL-15-producing natural killer cells in B-cell acute lymphoblastic leukemia
Abstract
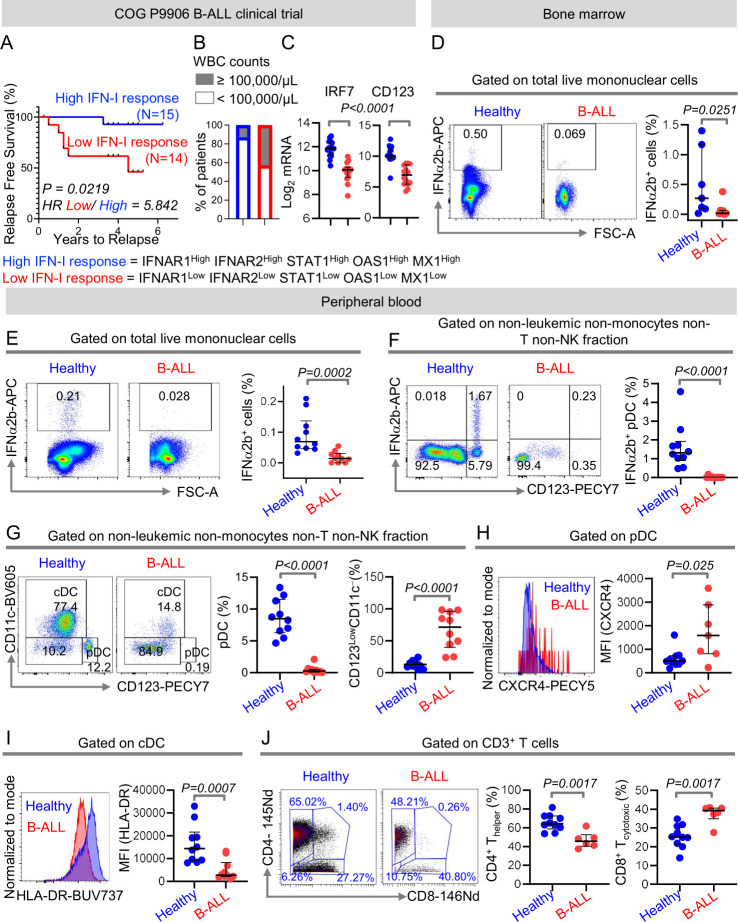
Background: Type I interferons (IFN-Is), secreted by hematopoietic cells, drive immune surveillance of solid tumors. However, the mechanisms of suppression of IFN-I-driven immune responses in hematopoietic malignancies including B-cell acute lymphoblastic leukemia (B-ALL) are unknown.
Methods: Using high-dimensional cytometry, we delineate the defects in IFN-I production and IFN-I-driven immune responses in high-grade primary human and mouse B-ALLs. We develop natural killer (NK) cells as therapies to counter the intrinsic suppression of IFN-I production in B-ALL.
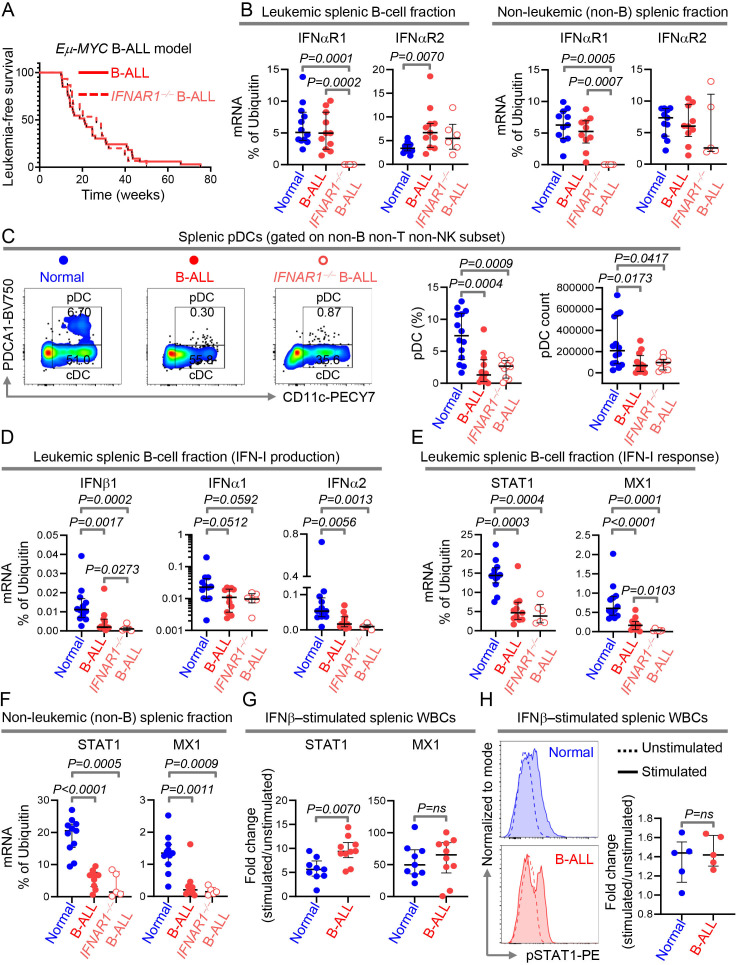
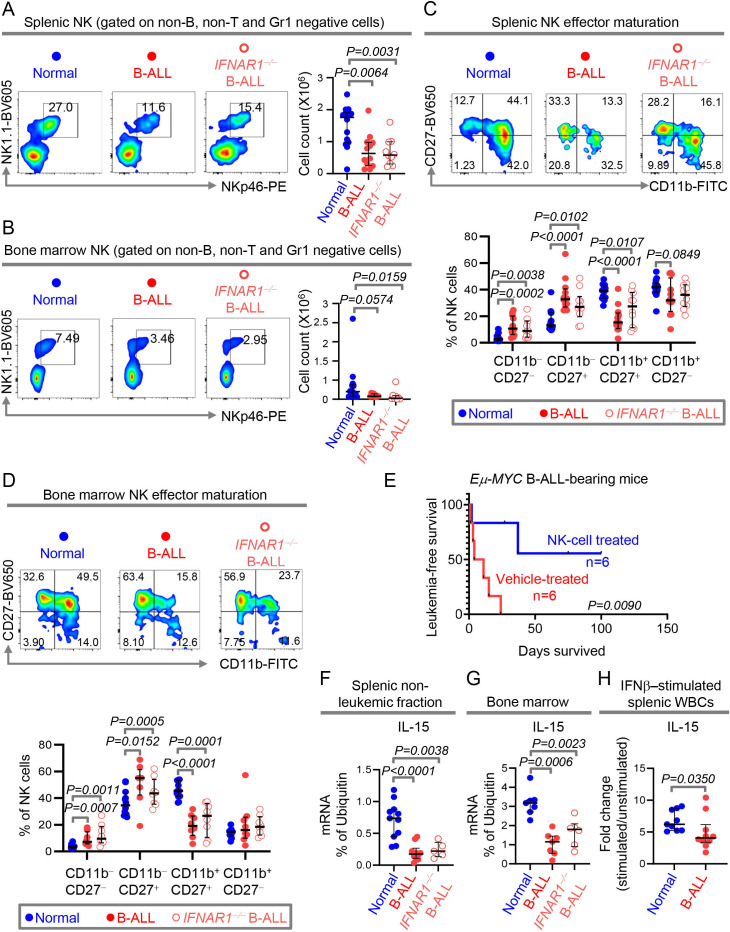
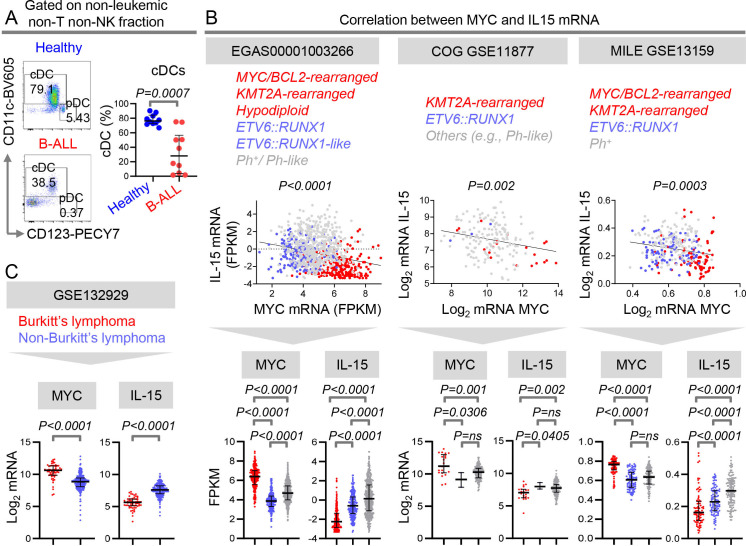
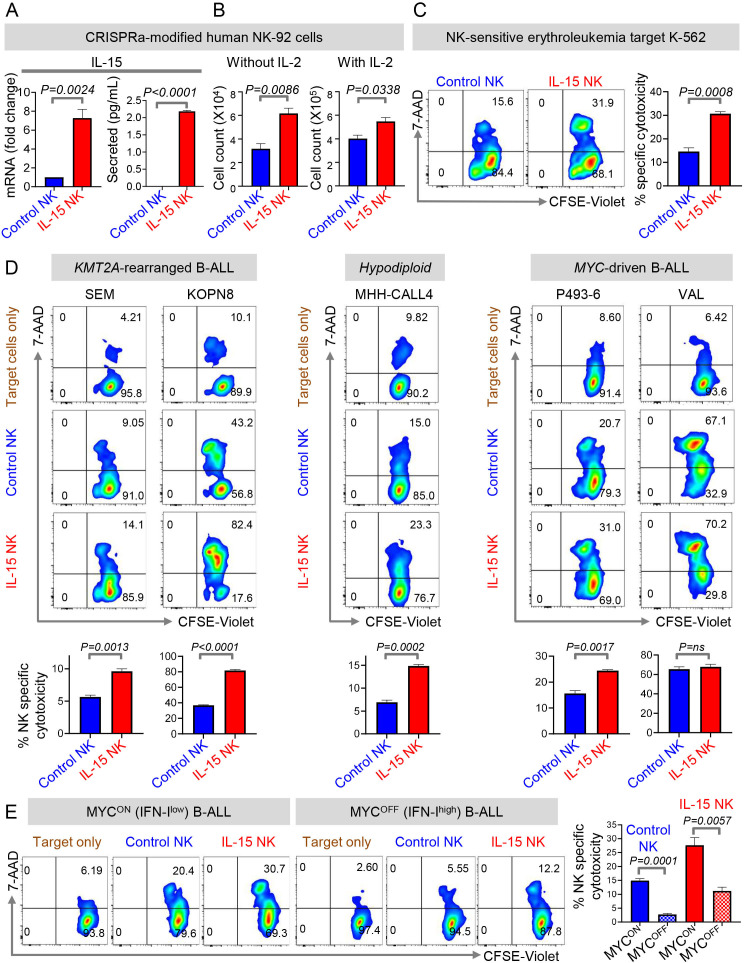
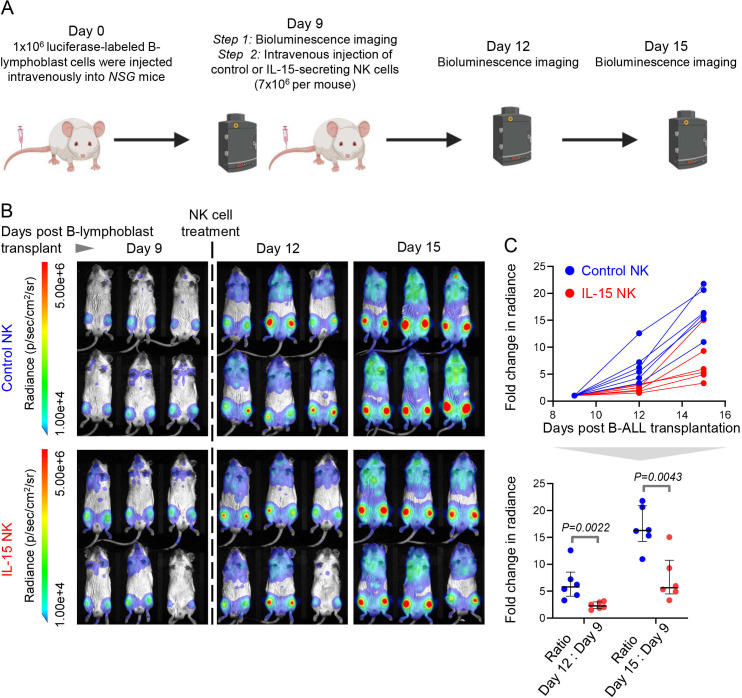
Results: We find that high expression of IFN-I signaling genes predicts favorable clinical outcome in patients with B-ALL, underscoring the importance of the IFN-I pathway in this malignancy. We show that human and mouse B-ALL microenvironments harbor an intrinsic defect in paracrine (plasmacytoid dendritic cell) and/or autocrine (B-cell) IFN-I production and IFN-I-driven immune responses. Reduced IFN-I production is sufficient for suppressing the immune system and promoting leukemia development in mice prone to MYC-driven B-ALL. Among anti-leukemia immune subsets, suppression of IFN-I production most markedly lowers the transcription of IL-15 and reduces NK-cell number and effector maturation in B-ALL microenvironments. Adoptive transfer of healthy NK cells significantly prolongs survival of overt ALL-bearing transgenic mice. Administration of IFN-Is to B-ALL-prone mice reduces leukemia progression and increases the frequencies of total NK and NK-cell effectors in circulation. Ex vivo treatment of malignant and non-malignant immune cells in primary mouse B-ALL microenvironments with IFN-Is fully restores proximal IFN-I signaling and partially restores IL-15 production. In B-ALL patients, the suppression of IL-15 is the most severe in difficult-to-treat subtypes with MYC overexpression. MYC overexpression promotes sensitivity of B-ALL to NK cell-mediated killing. To counter the suppressed IFN-I-induced IL-15 production in MYChigh human B-ALL, we CRISPRa-engineered a novel human NK-cell line that secretes IL-15. CRISPRa IL-15-secreting human NK cells kill high-grade human B-ALL in vitro and block leukemia progression in vivo more effectively than NK cells that do not produce IL-15.
Conclusion: We find that restoration of the intrinsically suppressed IFN-I production in B-ALL underlies the therapeutic efficacy of IL-15-producing NK cells and that such NK cells represent an attractive therapeutic solution for the problem of drugging MYC in high-grade B-ALL.
Keywords: cytokines; hematologic neoplasms; immune evation; immunologic surveillance; killer cells, natural.
© Author(s) (or their employer(s)) 2023. Re-use permitted under CC BY-NC. No commercial re-use. See rights and permissions. Published by BMJ.
Conflict of interest statement
Competing interests: The work described in this study is covered by pending US and PCT patent applications assigned to City of Hope and/or Stanford University with inventors SS, AK, ATK, CW-C, SJL, CD and HTM. MAC and JY are cofounders of CytoImmune Therapeutics.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources