

COX17 restricts renal fibrosis development by maintaining mitochondrial copper homeostasis and restoring complex IV activity
- PMID: 37217601
- PMCID: PMC10545728
- DOI: 10.1038/s41401-023-01098-3
COX17 restricts renal fibrosis development by maintaining mitochondrial copper homeostasis and restoring complex IV activity
Abstract
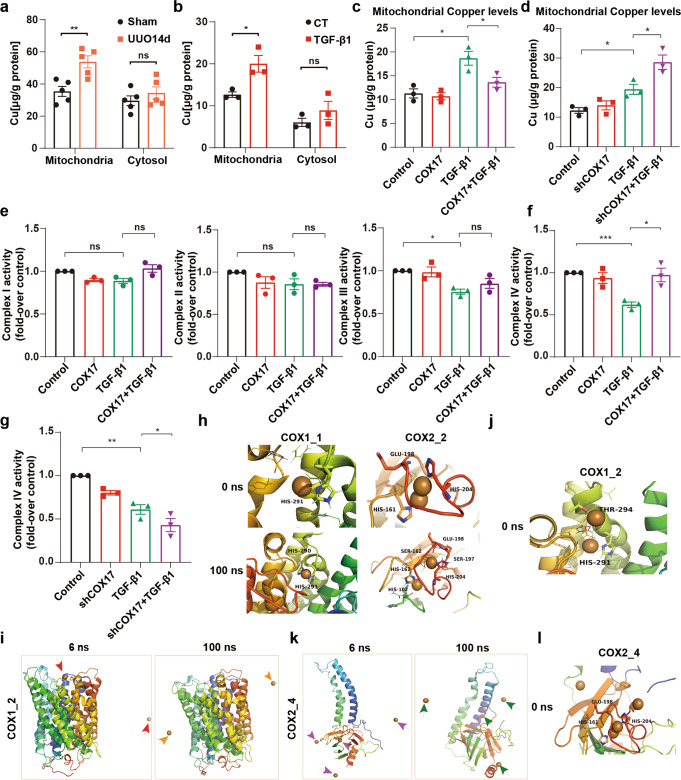
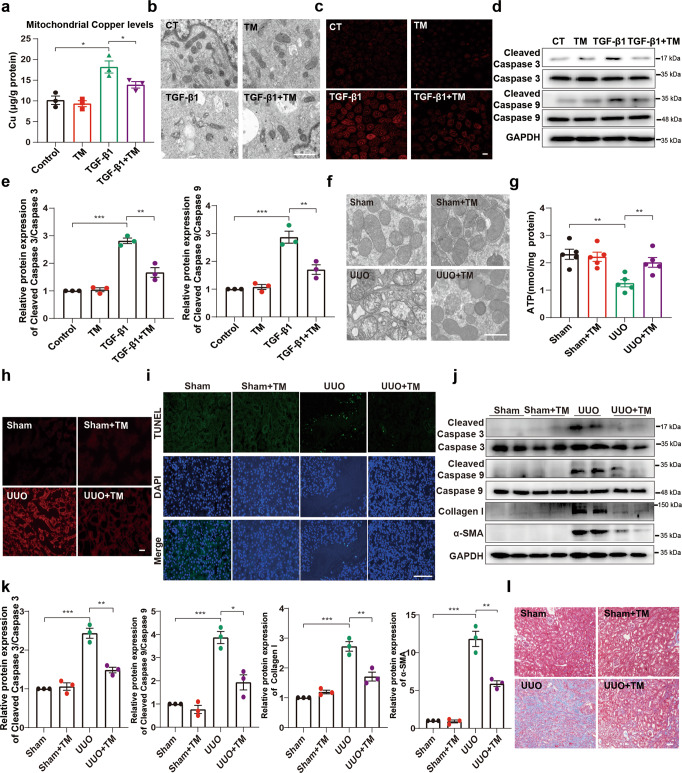
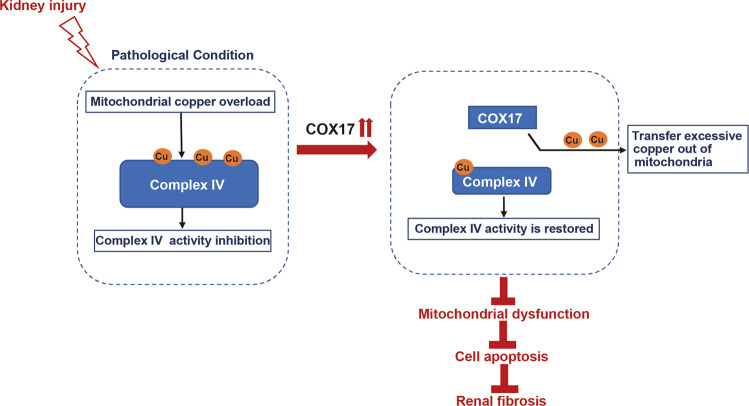
Renal fibrosis relies on multiple proteins and cofactors in its gradual development. Copper is a cofactor of many enzymes involved in renal microenvironment homeostasis. We previously reported that intracellular copper imbalance occurred during renal fibrosis development and was correlated with fibrosis intensity. In this study, we investigated the molecular mechanisms of how copper affected renal fibrosis development. Unilateral ureteral obstruction (UUO) mice were used for in vivo study; rat renal tubular epithelial cells (NRK-52E) treated with TGF-β1 were adapted as an in vitro fibrotic model. We revealed that the accumulation of copper in mitochondria, rather than cytosol, was responsible for mitochondrial dysfunction, cell apoptosis and renal fibrosis in both in vivo and in vitro fibrotic models. Furthermore, we showed that mitochondrial copper overload directly disrupted the activity of respiratory chain complex IV (cytochrome c oxidase), but not complex I, II and III, which hampered respiratory chain and disrupted mitochondrial functions, eventually leading to fibrosis development. Meanwhile, we showed that COX17, the copper chaperone protein, was significantly upregulated in the mitochondria of fibrotic kidneys and NRK-52E cells. Knockdown of COX17 aggravated mitochondrial copper accumulation, inhibited complex IV activity, augmented mitochondrial dysfunction and led to cell apoptosis and renal fibrosis, whereas overexpression of COX17 could discharge copper from mitochondria and protect mitochondrial function, alleviating renal fibrosis. In conclusion, copper accumulation in mitochondria blocks complex IV activity and induces mitochondrial dysfunction. COX17 plays a pivotal role in maintaining mitochondrial copper homeostasis, restoring complex IV activity, and ameliorating renal fibrosis.
Keywords: COX17; NRK-52E cells; copper; cytochrome c oxidase; mitochondria; renal fibrosis.
© 2023. The Author(s), under exclusive licence to Shanghai Institute of Materia Medica, Chinese Academy of Sciences and Chinese Pharmacological Society.
Conflict of interest statement
The authors declare no competing interests.
Figures
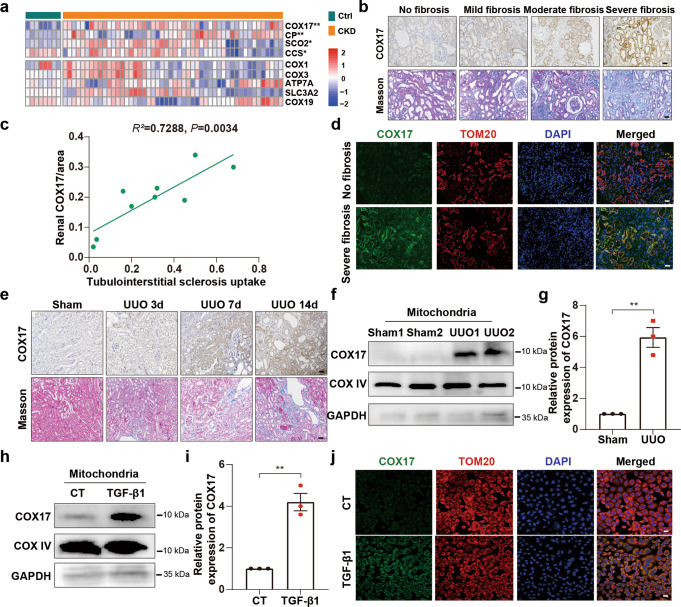
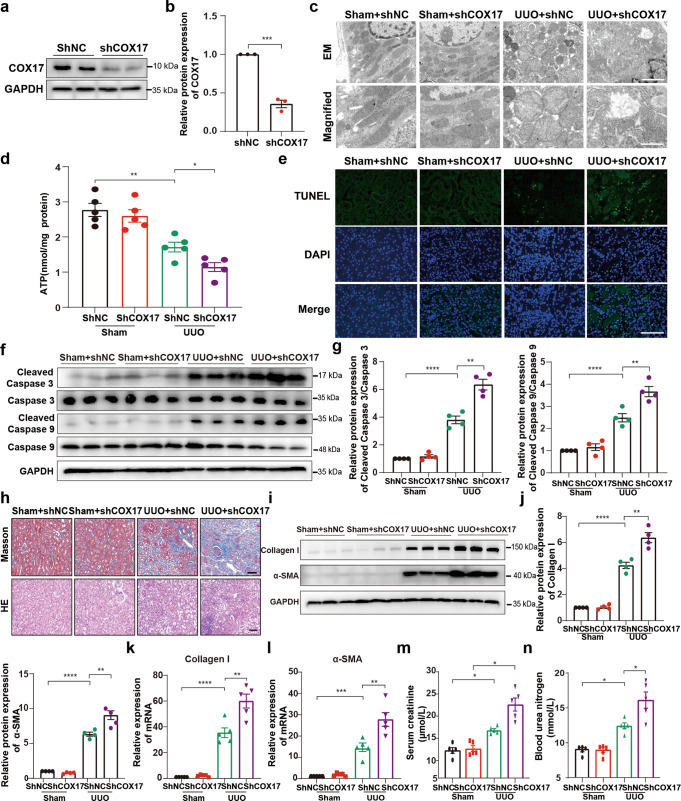
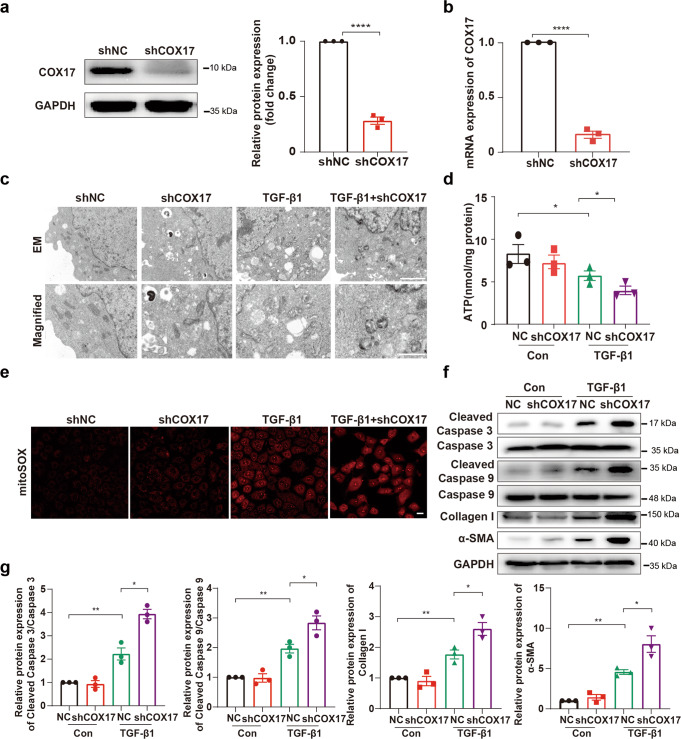
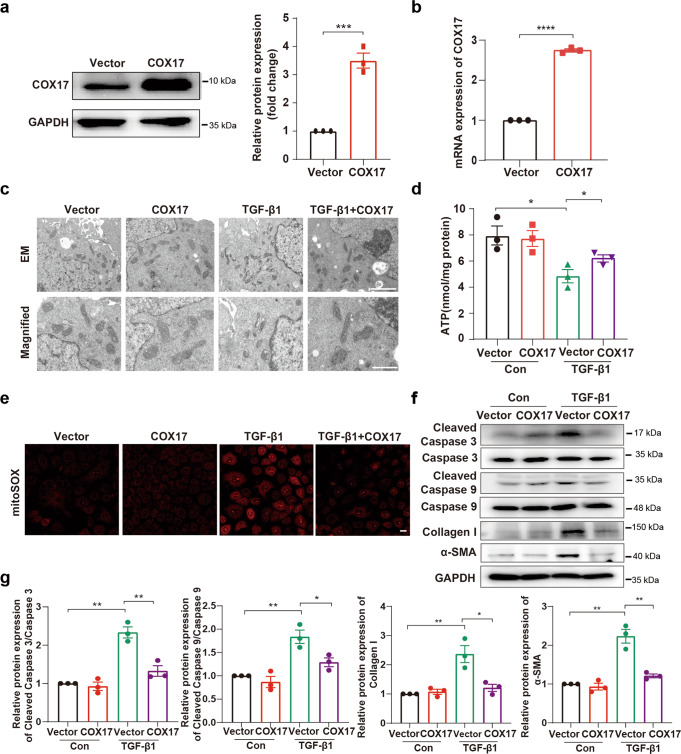
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
