

Synthetic Epigenetic Reprogramming of Mesenchymal to Epithelial States Using the CRISPR/dCas9 Platform in Triple Negative Breast Cancer
- PMID: 37217832
- PMCID: PMC10401103
- DOI: 10.1002/advs.202301802
Synthetic Epigenetic Reprogramming of Mesenchymal to Epithelial States Using the CRISPR/dCas9 Platform in Triple Negative Breast Cancer
Abstract
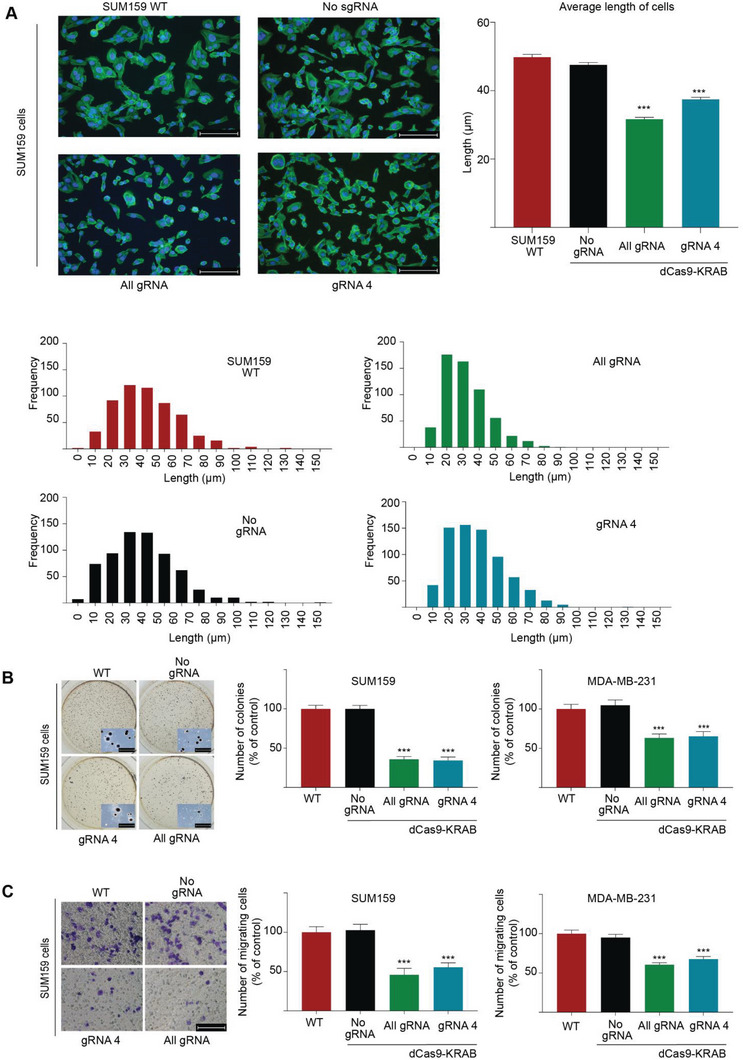
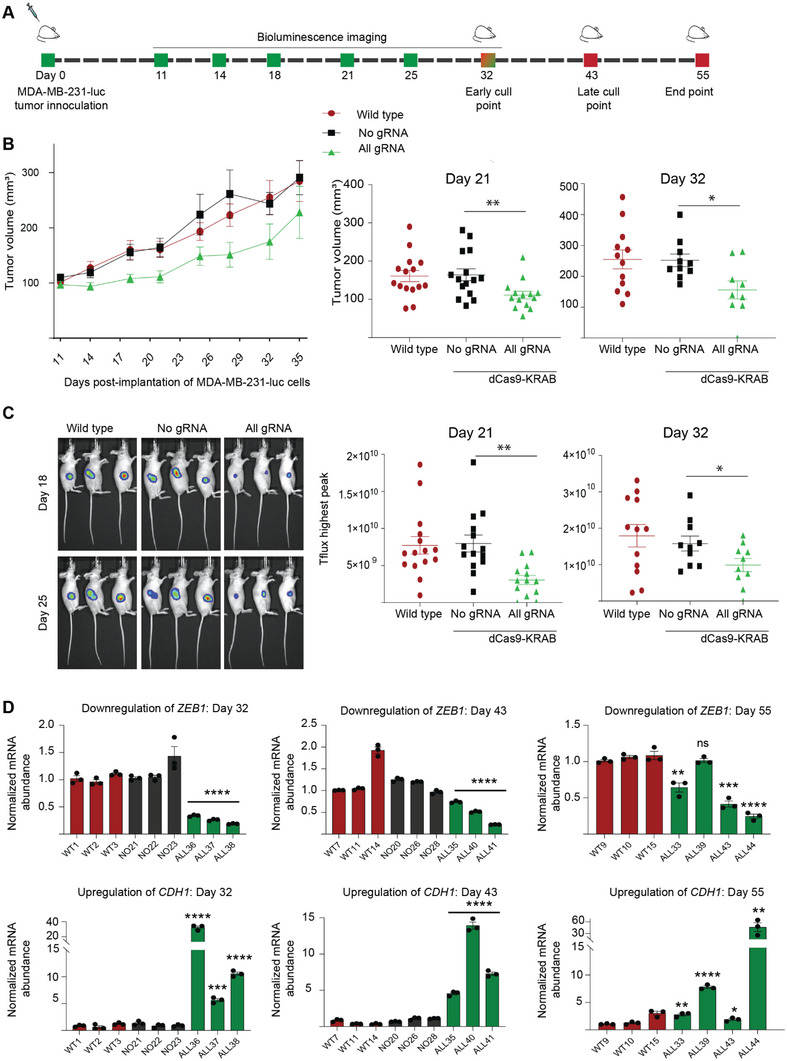
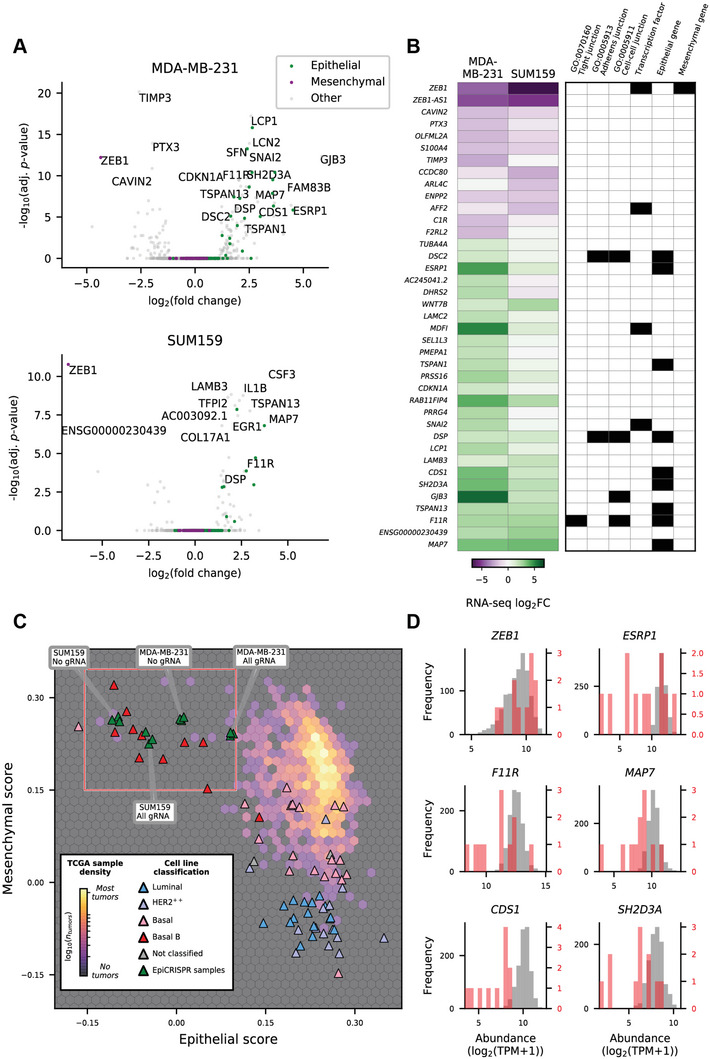
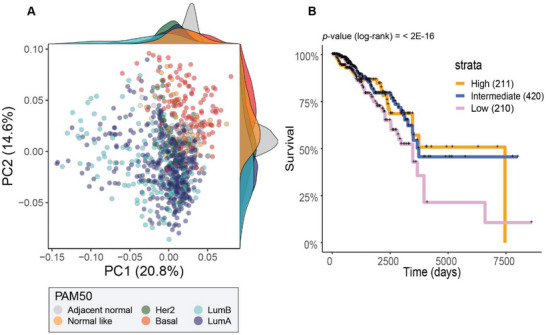
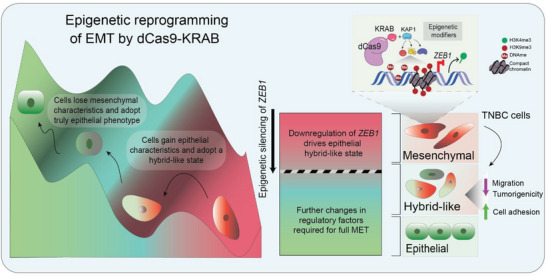
Epithelial-mesenchymal transition (EMT) is a reversible transcriptional program invoked by cancer cells to drive cancer progression. Transcription factor ZEB1 is a master regulator of EMT, driving disease recurrence in poor-outcome triple negative breast cancers (TNBCs). Here, this work silences ZEB1 in TNBC models by CRISPR/dCas9-mediated epigenetic editing, resulting in highly-specific and nearly complete suppression of ZEB1 in vivo, accompanied by long-lasting tumor inhibition. Integrated "omic" changes promoted by dCas9 linked to the KRAB domain (dCas9-KRAB) enabled the discovery of a ZEB1-dependent-signature of 26 genes differentially-expressed and -methylated, including the reactivation and enhanced chromatin accessibility in cell adhesion loci, outlining epigenetic reprogramming toward a more epithelial state. In the ZEB1 locus transcriptional silencing is associated with induction of locally-spread heterochromatin, significant changes in DNA methylation at specific CpGs, gain of H3K9me3, and a near complete erasure of H3K4me3 in the ZEB1 promoter. Epigenetic shifts induced by ZEB1-silencing are enriched in a subset of human breast tumors, illuminating a clinically-relevant hybrid-like state. Thus, the synthetic epi-silencing of ZEB1 induces stable "lock-in" epigenetic reprogramming of mesenchymal tumors associated with a distinct and stable epigenetic landscape. This work outlines epigenome-engineering approaches for reversing EMT and customizable precision molecular oncology approaches for targeting poor outcome breast cancers.
Keywords: CRISPR/dCas9 repression; ZEB1; cancer epigenetics; epithelial-mesenchymal transition; triple negative breast cancer.
© 2023 The Authors. Advanced Science published by Wiley-VCH GmbH.
Conflict of interest statement
The authors declare no conflict of interest.
Figures



References
-
- Yang J., Antin P., Berx G., Blanpain C., Brabletz T., Bronner M., Campbell K., Cano A., Casanova J., Christofori G., Dedhar S., Derynck R., Ford H. L., Fuxe J., García De Herreros A., Goodall G. J., Hadjantonakis A.‐K., Huang R. Y. J., Kalcheim C., Kalluri R., Kang Y., Khew‐Goodall Y., Levine H., Liu J., Longmore G. D., Mani S. A., Massagué J., Mayor R., Mcclay D., Mostov K. E., et al., Nat. Rev. Mol. Cell Biol. 2020, 21, 341. - PMC - PubMed
-
- Chaffer C. L., Thompson E. W., Williams E. D., Cells Tissues Organs 2007, 185, 7. - PubMed
-
- Brabletz T., Hlubek F., Spaderna S., Schmalhofer O., Hiendlmeyer E., Jung A., Kirchner T., Cells Tissues Organs 2005, 179, 56. - PubMed
-
- Jolly M. K., Murphy R. J., Bhatia S., Whitfield H. J., Redfern A., Davis M. J., Thompson E. W., Cells Tissues Organs 2022, 211, 110. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- APP1147528/National Health and Medical Research Council
- FT130101767/Australian Research Council Future Fellowship
- Cancer Council of Western Australia Research Fellowship
- APP1165208/Future Health Research and Innovation
- APP1187328/Future Health Research and Innovation
- IIRS-22-044/National Breast Cancer Foundation
- 19-009/CBCF-NBCF Brain and Breast Cancer Research Collaboration Initiative
- APP2013068/Cancer Council New South Wales
- APP2004608/Cancer Council Western Australia
- GNT1178460/Cancer Council Western Australia
- Howard Hughes Medical Institute International Research
- Australian Cancer Research Foundation Centre for Advanced Cancer Genomics and Genomics WA
- Harry Perkins Institute of Medical Research
- Australian Genome Research Facility
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials