

Physical exercise attenuates age-related muscle atrophy and exhibits anti-ageing effects via the adiponectin receptor 1 signalling
- PMID: 37222007
- PMCID: PMC10401527
- DOI: 10.1002/jcsm.13257
Physical exercise attenuates age-related muscle atrophy and exhibits anti-ageing effects via the adiponectin receptor 1 signalling
Abstract
Background: Although the adiponectin signalling exerts exercise-mimicking effects, whether this pathway contributes to the anti-ageing benefits of physical exercise has not been established yet.
Methods: Swim exercise training and wheel running were used to measure lifespan in the nematode Caenorhabditis elegans and skeletal muscle quality in mice, respectively. Muscle weight, muscle fibre cross-sectional area (CSA) and myonuclei number were used to evaluate muscle mass. RNA sequencing (RNA-Seq) analysis of skeletal muscle in exercised mice was used to study the underlying mechanisms. Western blot and immunofluorescence were performed to explore autophagy- and senescence-related markers.
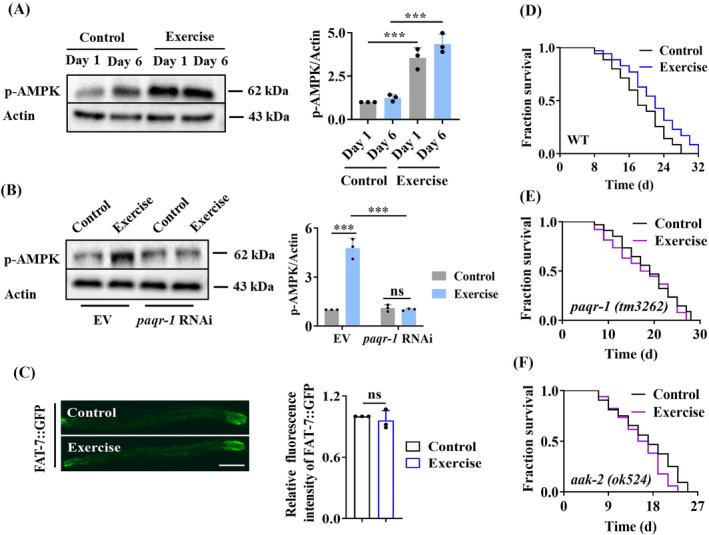
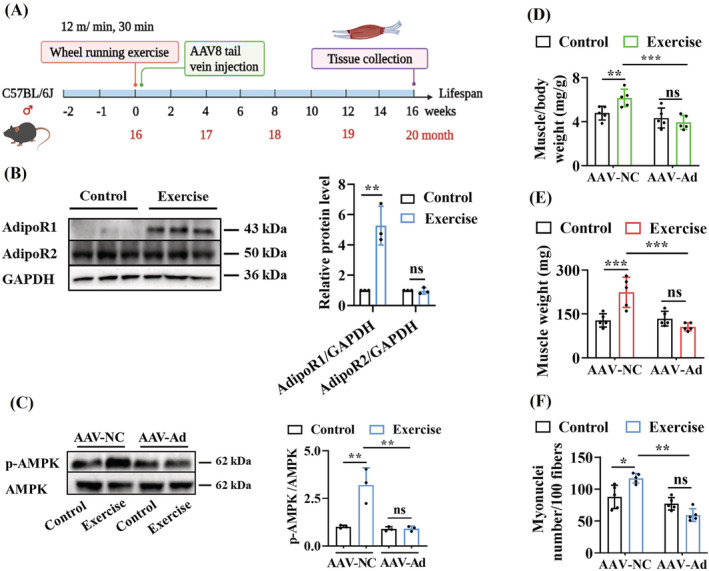
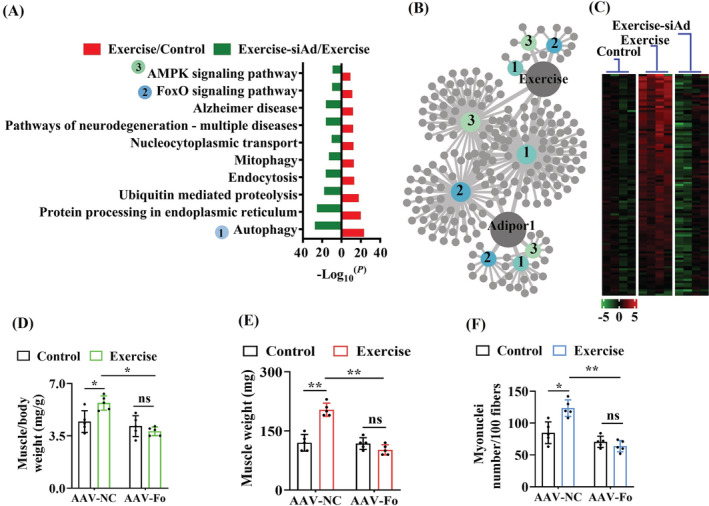
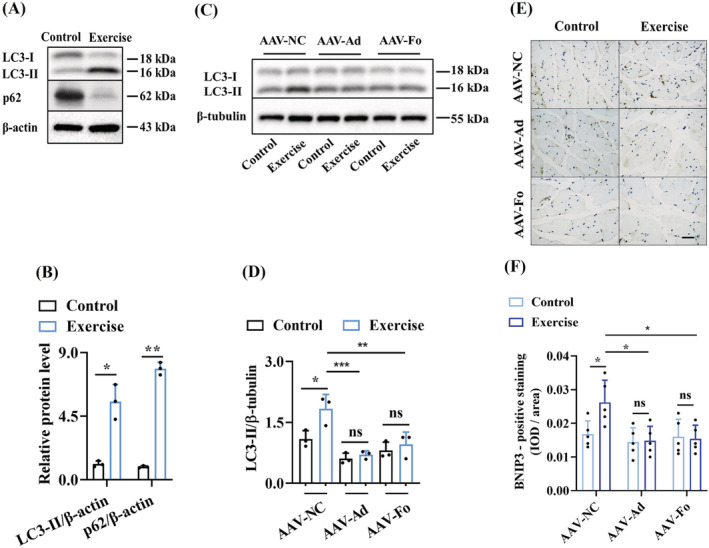
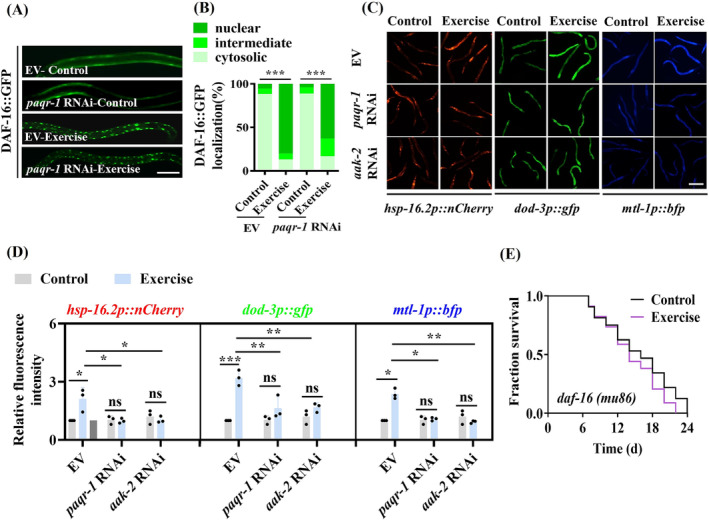
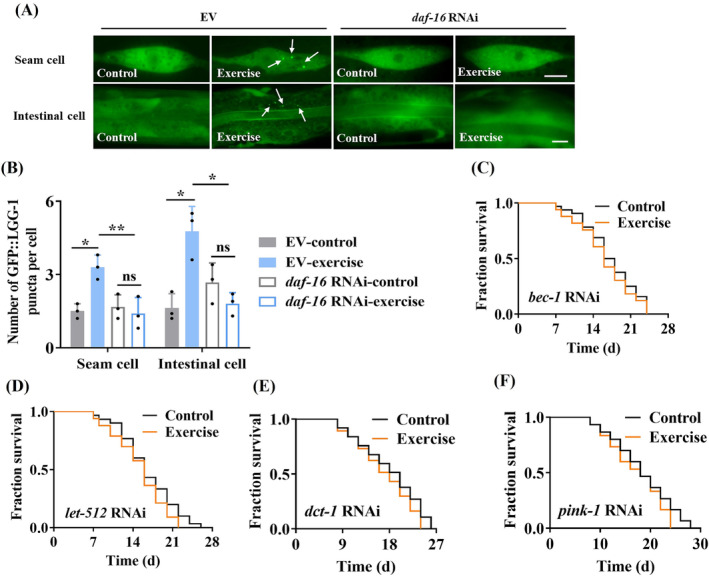
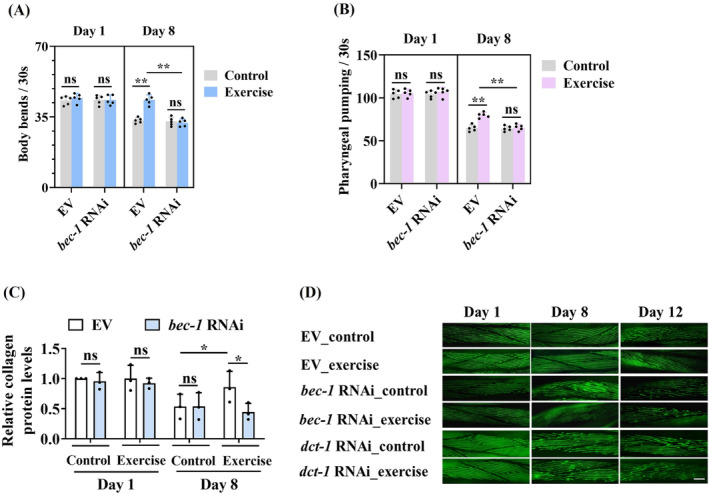
Results: The C. elegans adiponectin receptor PAQR-1/AdipoR1, but not PAQR-2/AdipoR2, was activated (3.55-fold and 3.48-fold increases in p-AMPK on Days 1 and 6, respectively, P < 0.001), which was involved in lifespan extension in exercised worms. Exercise training increased skeletal muscle mass index (1.29-fold, P < 0.01), muscle weight (1.75-fold, P < 0.001), myonuclei number (1.33-fold, P < 0.05), muscle fibre CSA (1.39-fold, P < 0.05) and capillary abundance (2.19-fold, P < 0.001 for capillary density; 1.58-fold, P < 0.01 for capillary number) in aged mice. Physical exercise reduced protein (2.94-fold, P < 0.001) and mRNA levels (1.70-fold, P < 0.001) of p16INK4a , a marker for cellular senescence, in skeletal muscle of aged mice. These beneficial effects of exercise on skeletal muscle of mice were dependent on AdipoR1. Kyoto Encyclopedia of Genes and Genomes (KEGG) analysis for differentially expressed genes in skeletal muscle between exercised mice with and without AdipoR1 knockdown by RNA-Seq analysis revealed that several KEGG pathways, such as 'AMPK signalling pathway' (P < 0.001), 'FOXO signalling pathway' (P < 0.001) and 'autophagy' (P < 0.001) were overrepresented. Knockdown of FoxO3a inhibited exercise-mediated beneficial effects on skeletal muscle quality of mice by inhibiting autophagy/mitophagy (3.81-fold reduction in LC3-II protein, P < 0.001; 1.53-fold reduction in BNIP3 protein, P < 0.05). Knockdown of daf-16, the FoxO homologue in C. elegans, reduced autophagy (2.77-fold and 2.06-fold reduction in GFP::LGG-1 puncta in seam cells and the intestine, respectively, P < 0.05) and blocked lifespan extension by exercise in worms.
Conclusions: Our findings provide insights into how the AdipoR1 pathway has an impact on the anti-ageing benefits of exercise and implicate that activation of the AdipoR1 signalling may represent a potential therapeutic strategy for reducing age-related loss of skeletal muscle.
Keywords: AdipoR1; C. elegans; FOXO; autophagy; exercise; rodent; skeletal muscle loss.
© 2023 The Authors. Journal of Cachexia, Sarcopenia and Muscle published by John Wiley & Sons Ltd on behalf of Society on Sarcopenia, Cachexia and Wasting Disorders.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures
References
-
- Watanabe LP, Riddle NC. New opportunities: Drosophila as a model system for exercise research. J Appl Physiol 2019;127:482–490. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
