

Memory Th17 cell-mediated protection against lethal secondary pneumococcal pneumonia following influenza infection
- PMID: 37222516
- PMCID: PMC10470593
- DOI: 10.1128/mbio.00519-23
Memory Th17 cell-mediated protection against lethal secondary pneumococcal pneumonia following influenza infection
Abstract
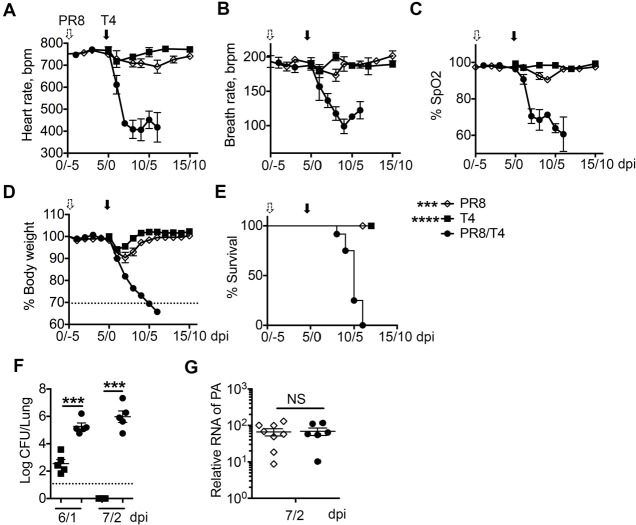
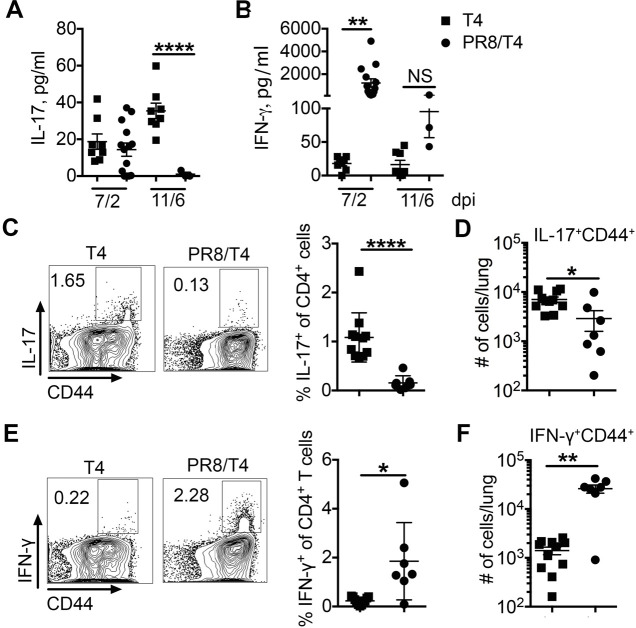
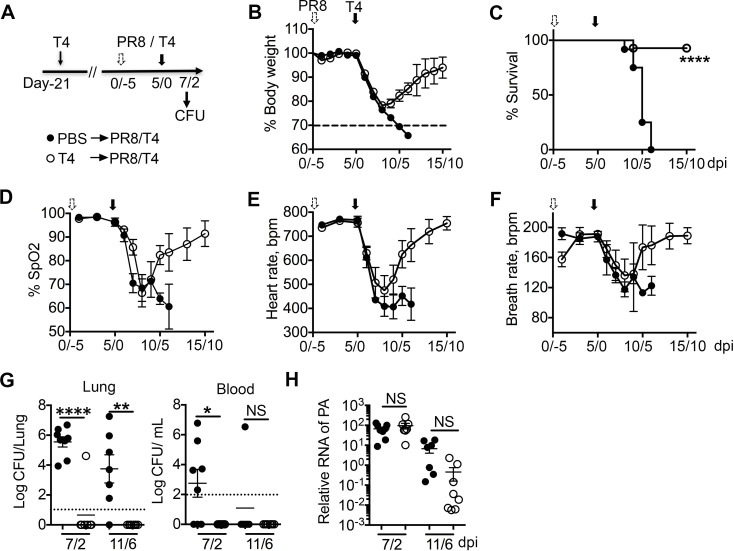
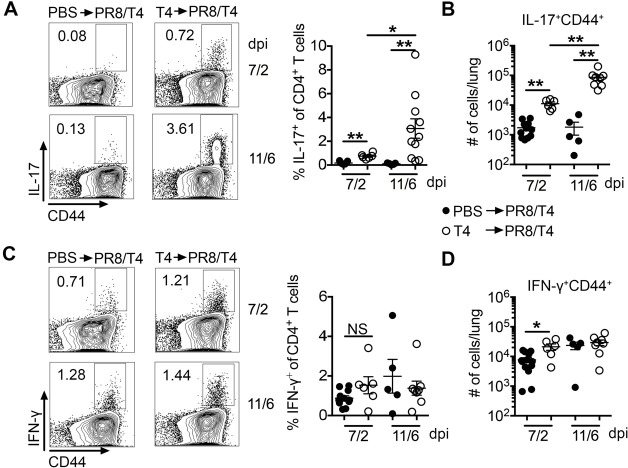
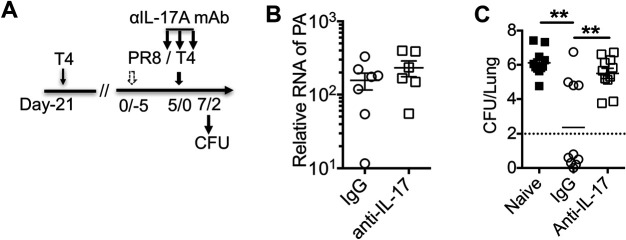
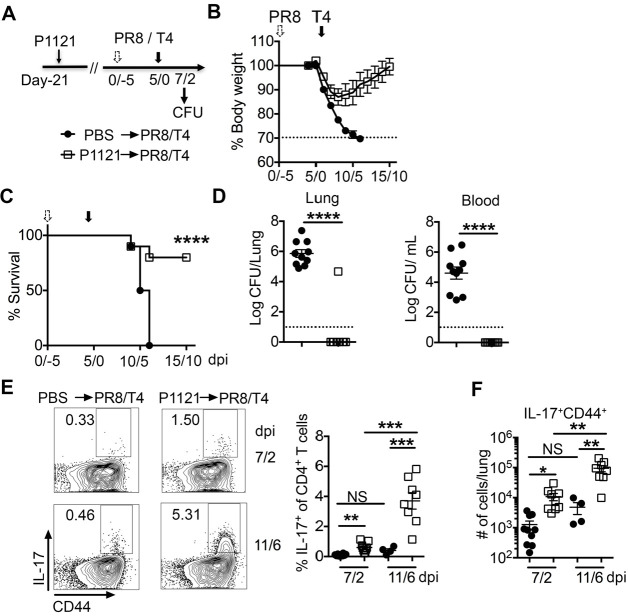
Streptococcus pneumoniae (Sp) frequently causes secondary pneumonia after influenza A virus (IAV) infection, leading to high morbidity and mortality worldwide. Concomitant pneumococcal and influenza vaccination improves protection against coinfection but does not always yield complete protection. Impaired innate and adaptive immune responses have been associated with attenuated bacterial clearance in influenza virus-infected hosts. In this study, we showed that preceding low-dose IAV infection caused persistent Sp infection and suppression of bacteria-specific T-helper type 17 (Th17) responses in mice. Prior Sp infection protected against subsequent IAV/Sp coinfection by improving bacterial clearance and rescuing bacteria-specific Th17 responses in the lungs. Furthermore, blockade of IL-17A by anti-IL-17A antibodies abrogated the protective effect of Sp preinfection. Importantly, memory Th17 responses induced by Sp preinfection overcame viral-driven Th17 inhibition and provided cross-protection against different Sp serotypes following coinfection with IAV. These results indicate that bacteria-specific Th17 memory cells play a key role in providing protection against IAV/Sp coinfection in a serotype-independent manner and suggest that a Th17-based vaccine would have excellent potential to mitigate disease caused by coinfection. IMPORTANCE Streptococcus pneumoniae (Sp) frequently causes secondary bacterial pneumonia after influenza A virus (IAV) infection, leading to increased morbidity and mortality worldwide. Current pneumococcal vaccines induce highly strain-specific antibody responses and provide limited protection against IAV/Sp coinfection. Th17 responses are broadly protective against Sp single infection, but whether the Th17 response, which is dramatically impaired by IAV infection in naïve mice, might be effective in immunization-induced protection against pneumonia caused by coinfection is not known. In this study, we have revealed that Sp-specific memory Th17 cells rescue IAV-driven inhibition and provide cross-protection against subsequent lethal coinfection with IAV and different Sp serotypes. These results indicate that a Th17-based vaccine would have excellent potential to mitigate disease caused by IAV/Sp coinfection.
Keywords: Streptococcus pneumoniae; Th17 responses; coinfection; cross-protection; influenza A virus.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Influenza A Virus Infection Predisposes Hosts to Secondary Infection with Different Streptococcus pneumoniae Serotypes with Similar Outcome but Serotype-Specific Manifestation.Infect Immun. 2016 Nov 18;84(12):3445-3457. doi: 10.1128/IAI.00422-16. Print 2016 Dec. Infect Immun. 2016. PMID: 27647871 Free PMC article.
-
Antigen-specific memory Th17 cells promote cross-protection against nontypeable Haemophilus influenzae after mild influenza A virus infection.Mucosal Immunol. 2023 Apr;16(2):153-166. doi: 10.1016/j.mucimm.2023.01.007. Epub 2023 Feb 2. Mucosal Immunol. 2023. PMID: 36736665 Free PMC article.
-
Bacterial factors required for Streptococcus pneumoniae coinfection with influenza A virus.J Biomed Sci. 2021 Aug 27;28(1):60. doi: 10.1186/s12929-021-00756-0. J Biomed Sci. 2021. PMID: 34452635 Free PMC article.
-
Modeling Influenza Virus Infection: A Roadmap for Influenza Research.Viruses. 2015 Oct 12;7(10):5274-304. doi: 10.3390/v7102875. Viruses. 2015. PMID: 26473911 Free PMC article. Review.
-
Secondary streptococcal infection following influenza.Microbiol Immunol. 2022 Jun;66(6):253-263. doi: 10.1111/1348-0421.12965. Epub 2022 May 26. Microbiol Immunol. 2022. PMID: 35088451 Review.
References
-
- Monsalvo AC, Batalle JP, Lopez MF, Krause JC, Klemenc J, Hernandez JZ, Maskin B, Bugna J, Rubinstein C, Aguilar L, Dalurzo L, Libster R, Savy V, Baumeister E, Aguilar L, Cabral G, Font J, Solari L, Weller KP, Johnson J, Echavarria M, Edwards KM, Chappell JD, Crowe JE, Williams JV, Melendi GA, Polack FP. 2011. Severe pandemic 2009 H1N1 influenza disease due to pathogenic immune complexes. Nat Med 17:195–199. doi:10.1038/nm.2262 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
