

PBP2b Mutations Improve the Growth of Phage-Resistant Lactococcus cremoris Lacking Polysaccharide Pellicle
- PMID: 37222606
- PMCID: PMC10304956
- DOI: 10.1128/aem.02103-22
PBP2b Mutations Improve the Growth of Phage-Resistant Lactococcus cremoris Lacking Polysaccharide Pellicle
Abstract
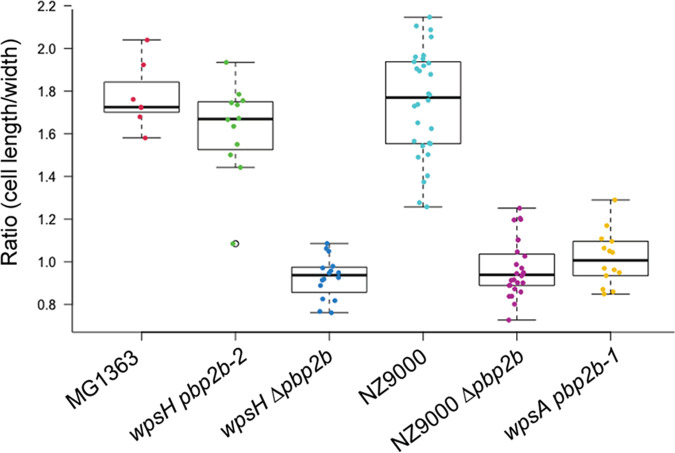
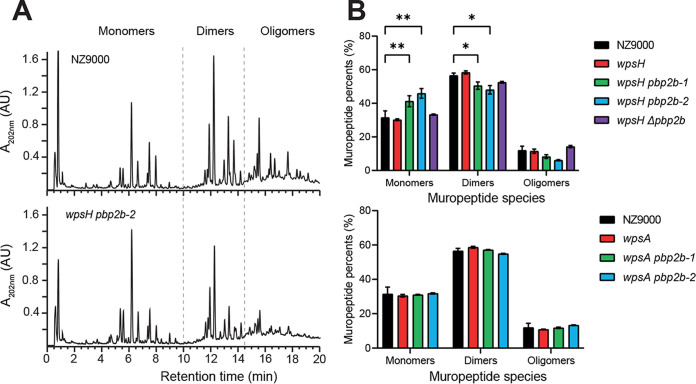
Lactococcus lactis and Lactococcus cremoris are Gram-positive lactic acid bacteria widely used as starter in milk fermentations. Lactococcal cells are covered with a polysaccharide pellicle (PSP) that was previously shown to act as the receptor for numerous bacteriophages of the Caudoviricetes class. Thus, mutant strains lacking PSP are phage resistant. However, because PSP is a key cell wall component, PSP-negative mutants exhibit dramatic alterations of cell shape and severe growth defects, which limit their technological value. In the present study, we isolated spontaneous mutants with improved growth, from L. cremoris PSP-negative mutants. These mutants grow at rates similar to the wild-type strain, and based on transmission electron microscopy analysis, they exhibit improved cell morphology compared to their parental PSP-negative mutants. In addition, the selected mutants maintain their phage resistance. Whole-genome sequencing of several such mutants showed that they carried a mutation in pbp2b, a gene encoding a penicillin-binding protein involved in peptidoglycan biosynthesis. Our results indicate that lowering or turning off PBP2b activity suppresses the requirement for PSP and ameliorates substantially bacterial fitness and morphology. IMPORTANCE Lactococcus lactis and Lactococcus cremoris are widely used in the dairy industry as a starter culture. As such, they are consistently challenged by bacteriophage infections which may result in reduced or failed milk acidification with associated economic losses. Bacteriophage infection starts with the recognition of a receptor at the cell surface, which was shown to be a cell wall polysaccharide (the polysaccharide pellicle [PSP]) for the majority of lactococcal phages. Lactococcal mutants devoid of PSP exhibit phage resistance but also reduced fitness, since their morphology and division are severely impaired. Here, we isolated spontaneous, food-grade non-PSP-producing L. cremoris mutants resistant to bacteriophage infection with a restored fitness. This study provides an approach to isolate non-GMO phage-resistant L. cremoris and L. lactis strains, which can be applied to strains with technological functionalities. Also, our results highlight for the first time the link between peptidoglycan and cell wall polysaccharide biosynthesis.
Keywords: Lactococcus; bacteriophage resistance; cell wall; lactic acid bacteria; penicillin-binding proteins; polysaccharide pellicle.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
MeSH terms
Substances
Supplementary concepts
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
