

The composition and function of Enterococcus faecalis membrane vesicles
- PMID: 37223255
- PMCID: PMC10117786
- DOI: 10.1093/femsml/uqab002
The composition and function of Enterococcus faecalis membrane vesicles
Abstract
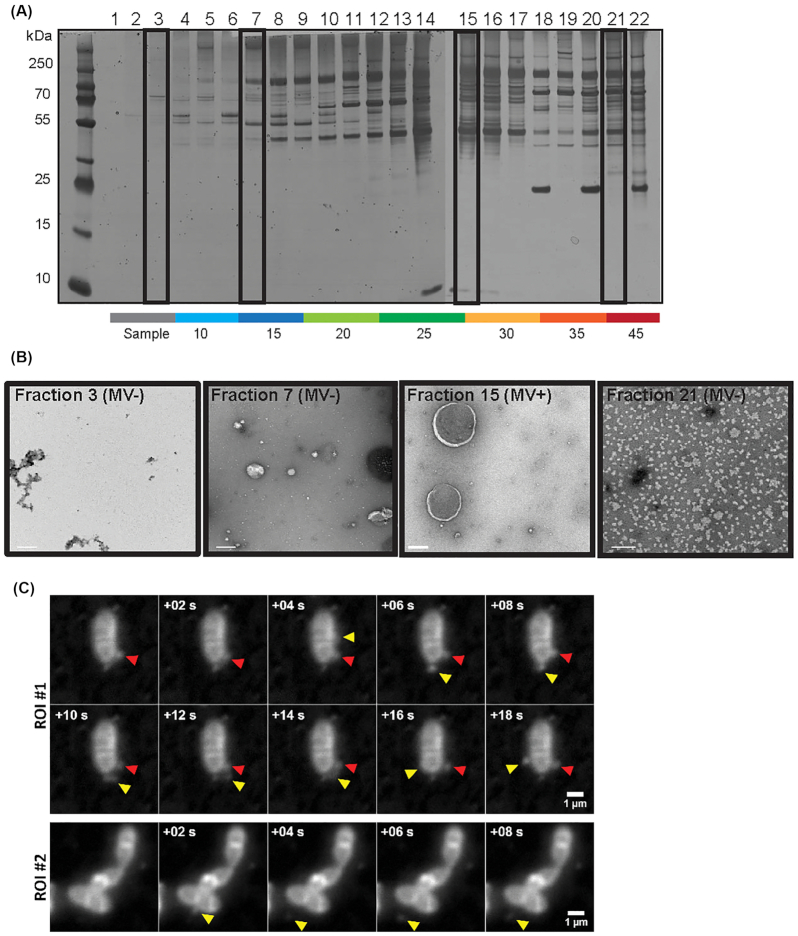
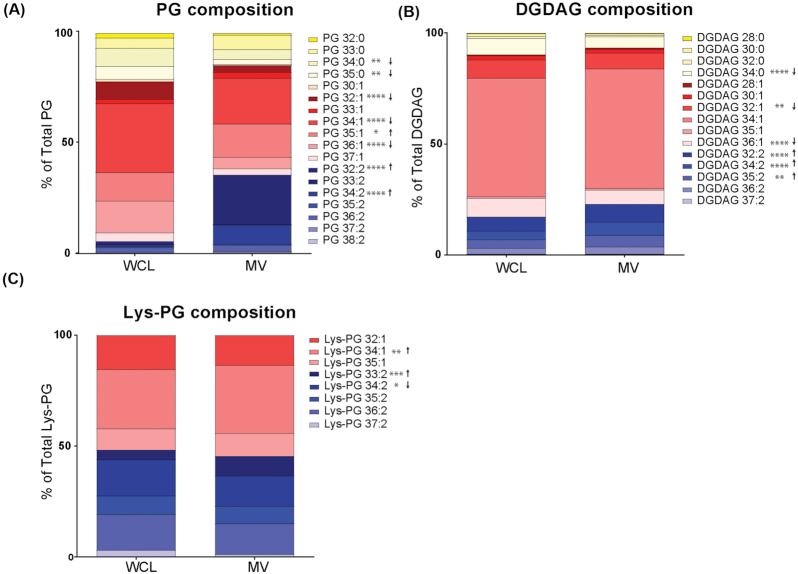
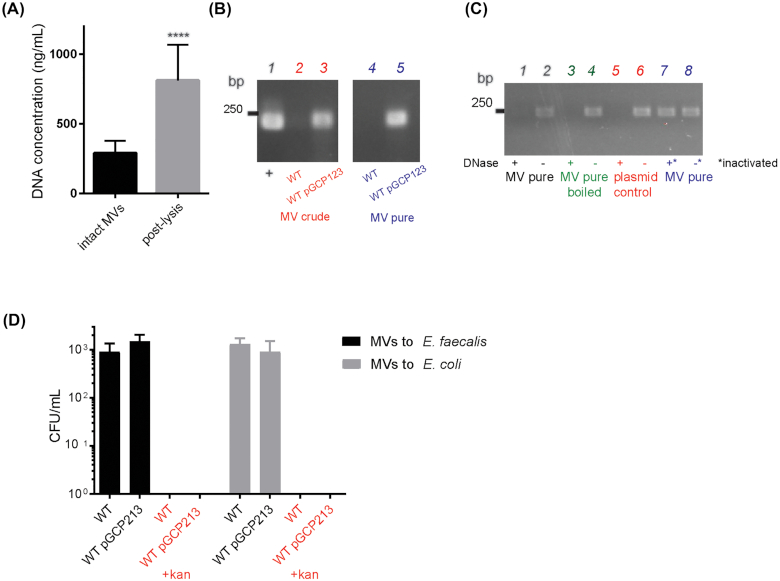
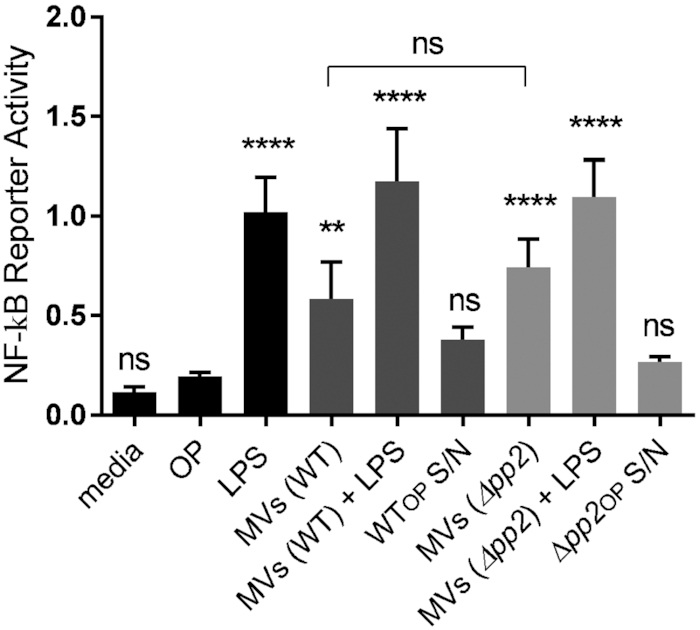
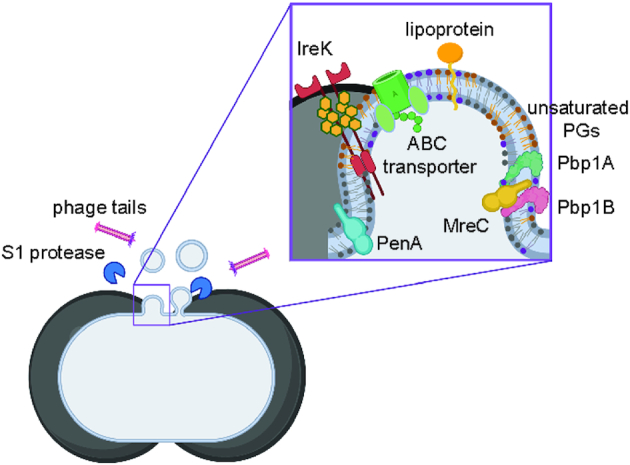
Membrane vesicles (MVs) contribute to various biological processes in bacteria, including virulence factor delivery, antimicrobial resistance, host immune evasion and cross-species communication. MVs are frequently released from the surface of both Gram-negative and Gram-positive bacteria during growth. In some Gram-positive bacteria, genes affecting MV biogenesis have been identified, but the mechanism of MV formation is unknown. In Enterococcus faecalis, a causative agent of life-threatening bacteraemia and endocarditis, neither mechanisms of MV formation nor their role in virulence has been examined. Since MVs of many bacterial species are implicated in host-pathogen interactions, biofilm formation, horizontal gene transfer, and virulence factor secretion in other species, we sought to identify, describe and functionally characterize MVs from E. faecalis. Here, we show that E. faecalis releases MVs that possess unique lipid and protein profiles, distinct from the intact cell membrane and are enriched in lipoproteins. MVs of E. faecalis are specifically enriched in unsaturated lipids that might provide membrane flexibility to enable MV formation, providing the first insights into the mechanism of MV formation in this Gram-positive organism.
Keywords: Enterococcus faecalis; NF-kB signaling; horizontal gene transfer; lipidomics; membrane vesicles; proteomics.
© The Author(s) 2021. Published by Oxford University Press on behalf of FEMS.
Conflict of interest statement
None declared.
Figures

References
-
- Bligh EG, Dyer WJ. A rapid method of total lipid extraction and purification. Can J Biochem Physiol. 1959;37:911– 7. - PubMed
LinkOut - more resources
Full Text Sources