

Revealing the small proteome of Haloferax volcanii by combining ribosome profiling and small-protein optimized mass spectrometry
- PMID: 37223747
- PMCID: PMC10117724
- DOI: 10.1093/femsml/uqad001
Revealing the small proteome of Haloferax volcanii by combining ribosome profiling and small-protein optimized mass spectrometry
Abstract
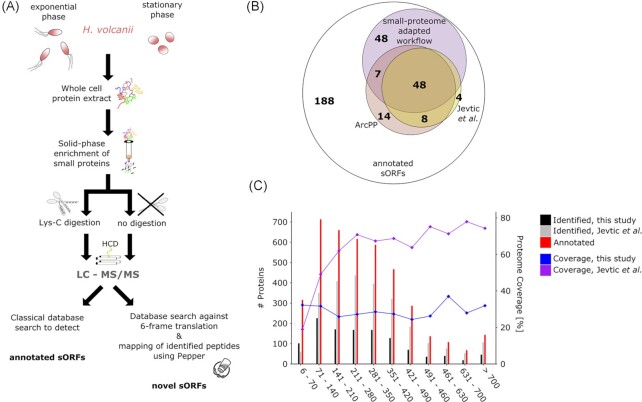
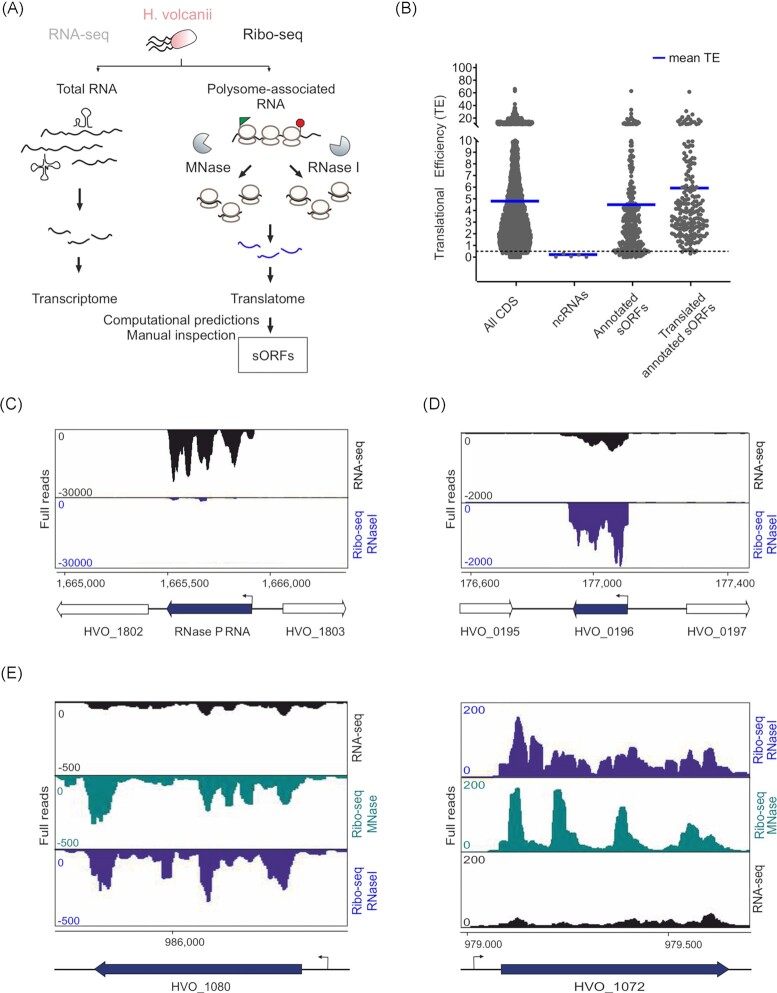
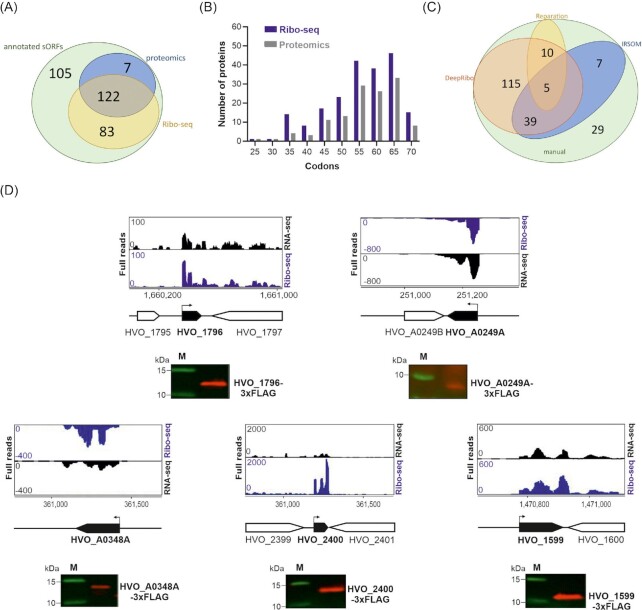
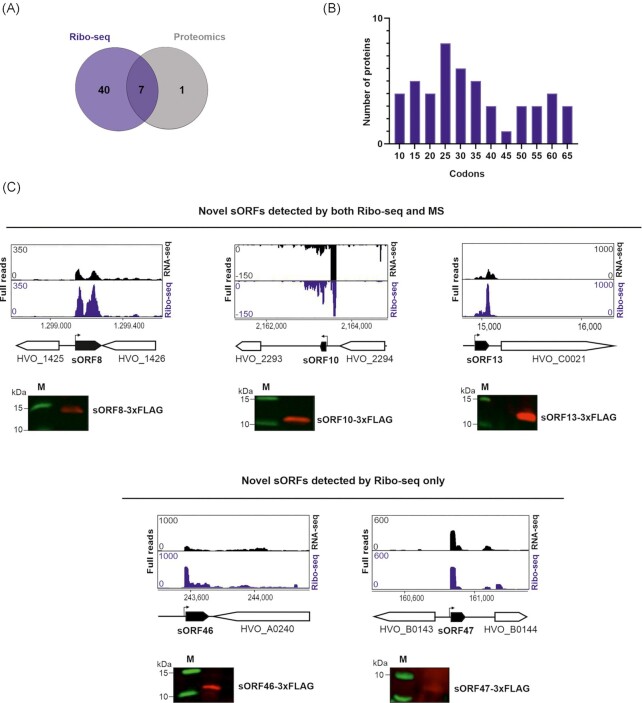
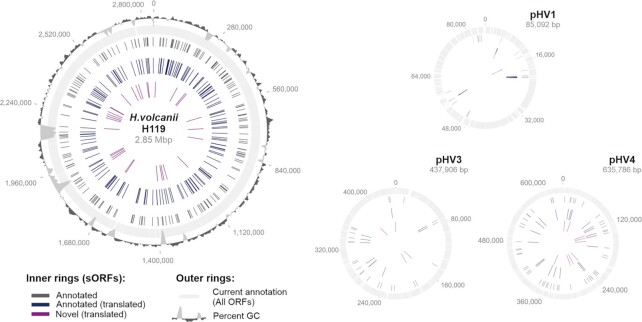
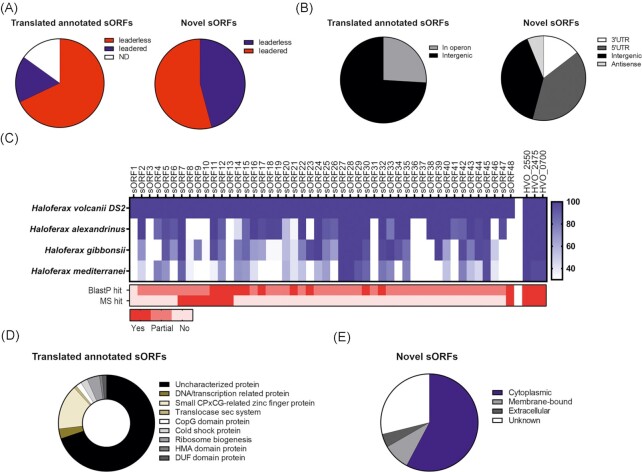
In contrast to extensively studied prokaryotic 'small' transcriptomes (encompassing all small noncoding RNAs), small proteomes (here defined as including proteins ≤70 aa) are only now entering the limelight. The absence of a complete small protein catalogue in most prokaryotes precludes our understanding of how these molecules affect physiology. So far, archaeal genomes have not yet been analyzed broadly with a dedicated focus on small proteins. Here, we present a combinatorial approach, integrating experimental data from small protein-optimized mass spectrometry (MS) and ribosome profiling (Ribo-seq), to generate a high confidence inventory of small proteins in the model archaeon Haloferax volcanii. We demonstrate by MS and Ribo-seq that 67% of the 317 annotated small open reading frames (sORFs) are translated under standard growth conditions. Furthermore, annotation-independent analysis of Ribo-seq data showed ribosomal engagement for 47 novel sORFs in intergenic regions. A total of seven of these were also detected by proteomics, in addition to an eighth novel small protein solely identified by MS. We also provide independent experimental evidence in vivo for the translation of 12 sORFs (annotated and novel) using epitope tagging and western blotting, underlining the validity of our identification scheme. Several novel sORFs are conserved in Haloferax species and might have important functions. Based on our findings, we conclude that the small proteome of H. volcanii is larger than previously appreciated, and that combining MS with Ribo-seq is a powerful approach for the discovery of novel small protein coding genes in archaea.
Keywords: Haloferax volcanii; Ribo-seq; archaea; mass spectrometry; proteomics; ribosome profiling; sORF; small protein; small proteome; sprotein.
© The Author(s) 2023. Published by Oxford University Press on behalf of FEMS.
Conflict of interest statement
None declared.
Figures
Similar articles
-
Unraveling the small proteome of the plant symbiont Sinorhizobium meliloti by ribosome profiling and proteogenomics.Microlife. 2023 Mar 10;4:uqad012. doi: 10.1093/femsml/uqad012. eCollection 2023. Microlife. 2023. PMID: 37223733 Free PMC article.
-
An update on sORFs.org: a repository of small ORFs identified by ribosome profiling.Nucleic Acids Res. 2018 Jan 4;46(D1):D497-D502. doi: 10.1093/nar/gkx1130. Nucleic Acids Res. 2018. PMID: 29140531 Free PMC article.
-
Genome-wide identification of transcriptional start sites in the haloarchaeon Haloferax volcanii based on differential RNA-Seq (dRNA-Seq).BMC Genomics. 2016 Aug 12;17(1):629. doi: 10.1186/s12864-016-2920-y. BMC Genomics. 2016. PMID: 27519343 Free PMC article.
-
Short open reading frames (sORFs) and microproteins: an update on their identification and validation measures.J Biomed Sci. 2022 Mar 17;29(1):19. doi: 10.1186/s12929-022-00802-5. J Biomed Sci. 2022. PMID: 35300685 Free PMC article. Review.
-
The power of the small: the underestimated role of small proteins in bacterial and archaeal physiology.Curr Opin Microbiol. 2023 Dec;76:102384. doi: 10.1016/j.mib.2023.102384. Epub 2023 Sep 28. Curr Opin Microbiol. 2023. PMID: 37776678 Review.
Cited by
-
Uncovering the small proteome of Methanosarcina mazei using Ribo-seq and peptidomics under different nitrogen conditions.Nat Commun. 2024 Oct 6;15(1):8659. doi: 10.1038/s41467-024-53008-8. Nat Commun. 2024. PMID: 39370430 Free PMC article.
-
Provirus deletion from Haloferax volcanii affects motility, stress resistance, and CRISPR RNA expression.Microlife. 2025 May 19;6:uqaf008. doi: 10.1093/femsml/uqaf008. eCollection 2025. Microlife. 2025. PMID: 40395997 Free PMC article.
-
Ribosome profiling reveals the fine-tuned response of Escherichia coli to mild and severe acid stress.mSystems. 2023 Dec 21;8(6):e0103723. doi: 10.1128/msystems.01037-23. Epub 2023 Nov 1. mSystems. 2023. PMID: 37909716 Free PMC article.
-
Characterization of the zinc finger μ-protein HVO_0758 from Haloferax volcanii: biological roles, zinc binding, and NMR solution structure.Front Microbiol. 2023 Nov 29;14:1280972. doi: 10.3389/fmicb.2023.1280972. eCollection 2023. Front Microbiol. 2023. PMID: 38094630 Free PMC article.
-
Investigation of the global translational response to oxidative stress in the model archaeon Haloferax volcanii reveals untranslated small RNAs with ribosome occupancy.bioRxiv [Preprint]. 2025 Jul 13:2025.04.08.647799. doi: 10.1101/2025.04.08.647799. bioRxiv. 2025. PMID: 40672279 Free PMC article. Preprint.
References
LinkOut - more resources
Full Text Sources
Molecular Biology Databases