

Development of new mutant alleles and markers for KTI1 and KTI3 via CRISPR/Cas9-mediated mutagenesis to reduce trypsin inhibitor content and activity in soybean seeds
- PMID: 37223818
- PMCID: PMC10200896
- DOI: 10.3389/fpls.2023.1111680
Development of new mutant alleles and markers for KTI1 and KTI3 via CRISPR/Cas9-mediated mutagenesis to reduce trypsin inhibitor content and activity in soybean seeds
Abstract
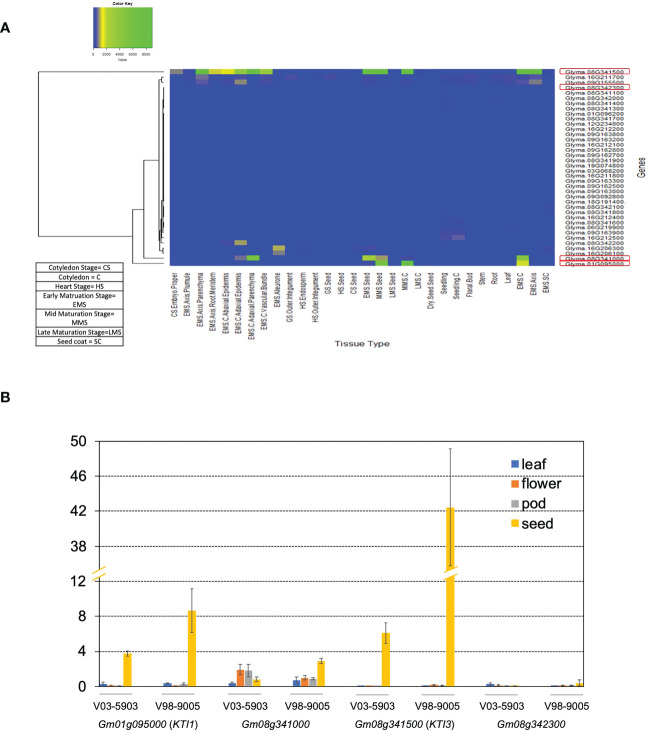
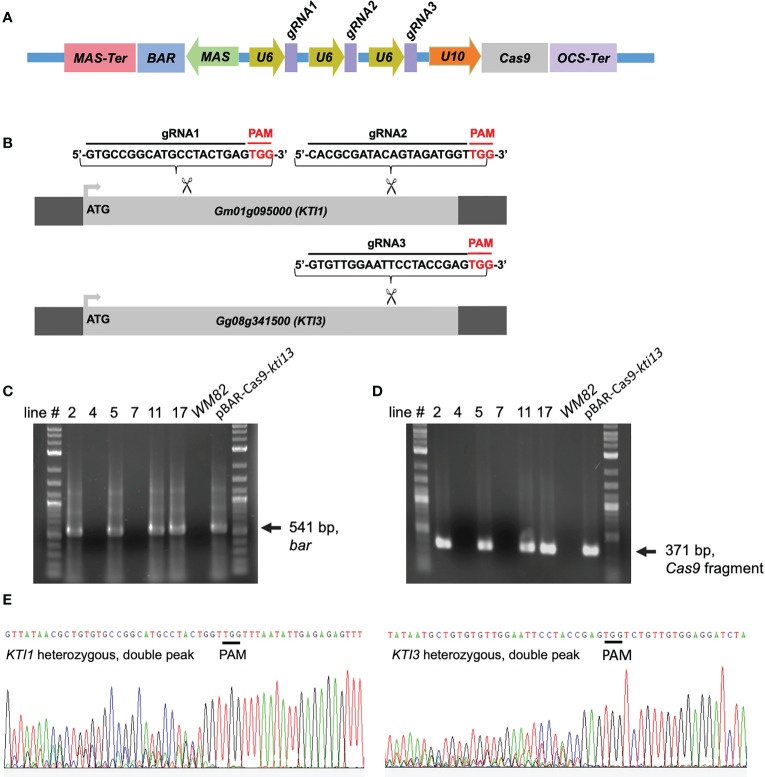
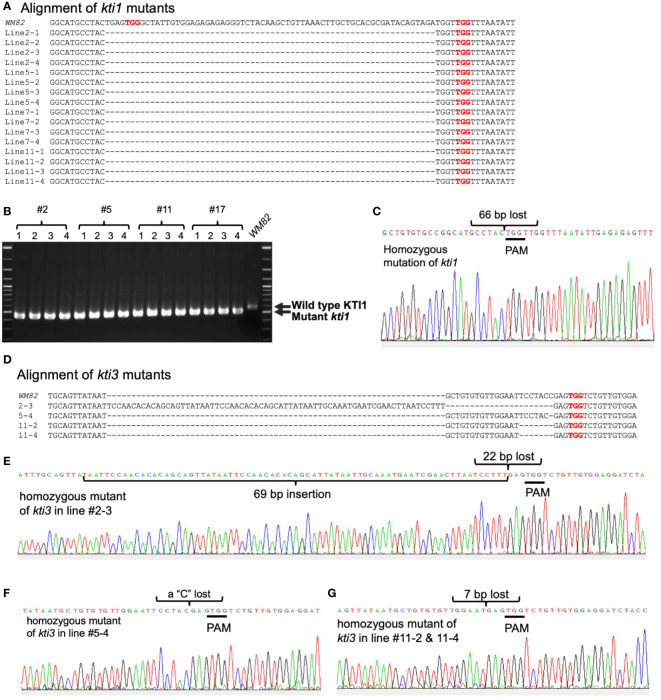
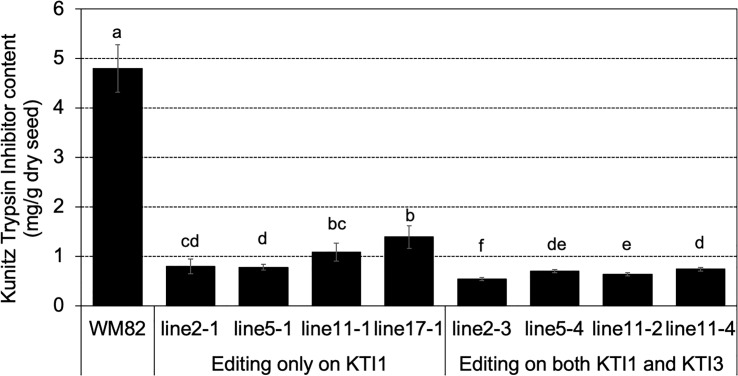
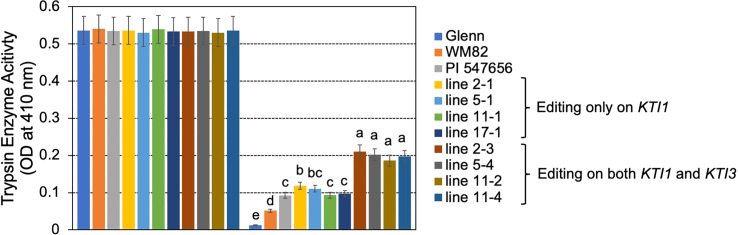
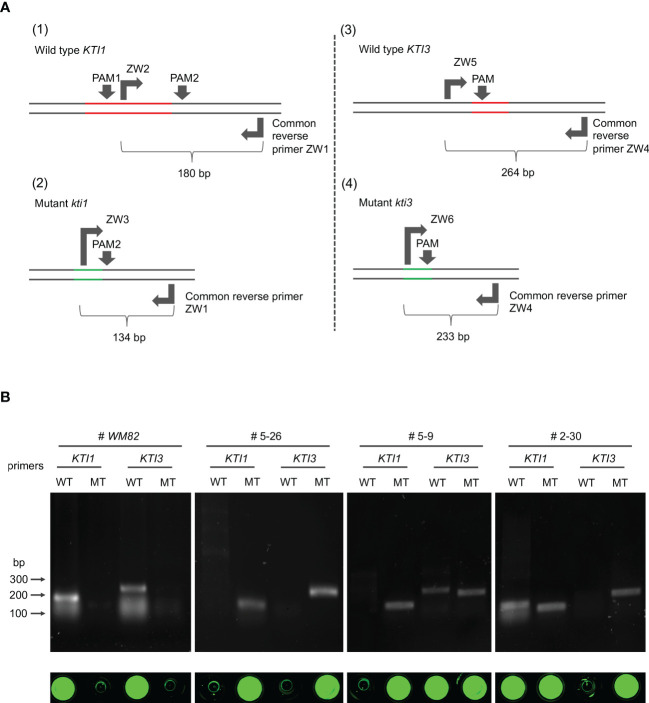
The digestibility of soybean meal can be severely impacted by trypsin inhibitor (TI), one of the most abundant anti-nutritional factors present in soybean seeds. TI can restrain the function of trypsin, a critical enzyme that breaks down proteins in the digestive tract. Soybean accessions with low TI content have been identified. However, it is challenging to breed the low TI trait into elite cultivars due to a lack of molecular markers associated with low TI traits. We identified Kunitz trypsin inhibitor 1 (KTI1, Gm01g095000) and KTI3 (Gm08g341500) as two seed-specific TI genes. Mutant kti1 and kti3 alleles carrying small deletions or insertions within the gene open reading frames were created in the soybean cultivar Glycine max cv. Williams 82 (WM82) using the CRISPR/Cas9-mediated genome editing approach. The KTI content and TI activity both remarkably reduced in kti1/3 mutants compared to the WM82 seeds. There was no significant difference in terms of plant growth or maturity days of kti1/3 transgenic and WM82 plants in greenhouse condition. We further identified a T1 line, #5-26, that carried double homozygous kti1/3 mutant alleles, but not the Cas9 transgene. Based on the sequences of kti1/3 mutant alleles in #5-26, we developed markers to co-select for these mutant alleles by using a gel-electrophoresis-free method. The kti1/3 mutant soybean line and associated selection markers will assist in accelerating the introduction of low TI trait into elite soybean cultivars in the future.
Keywords: CRISPR/Cas9; KTI1; KTI3; Kunitz trypsin inhibitor (KTI); allele-based selection marker; anti-nutritional factor; soybean.
Copyright © 2023 Wang, Shea, Rosso, Shang, Li, Bewick, Li, Zhao and Zhang.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
Similar articles
-
Validation of Molecular Markers for Low Kunitz Trypsin Inhibitor Content in European Soybean (Glycine max L. Merr.) Germplasm.Genes (Basel). 2024 Aug 5;15(8):1028. doi: 10.3390/genes15081028. Genes (Basel). 2024. PMID: 39202388 Free PMC article.
-
Overexpression of soybean trypsin inhibitor genes decreases defoliation by corn earworm (Helicoverpa zea) in soybean (Glycine max) and Arabidopsis thaliana.Front Plant Sci. 2023 Feb 17;14:1129454. doi: 10.3389/fpls.2023.1129454. eCollection 2023. Front Plant Sci. 2023. PMID: 36875574 Free PMC article.
-
Development of Breeder-Friendly KASP Markers for Low Concentration of Kunitz Trypsin Inhibitor in Soybean Seeds.Int J Mol Sci. 2021 Mar 6;22(5):2675. doi: 10.3390/ijms22052675. Int J Mol Sci. 2021. PMID: 33800919 Free PMC article.
-
Genomic introgression and expression profiling of the KTI null allele in soybean through elite-by-elite backcrossing.Plant Physiol Biochem. 2025 Jul;224:109912. doi: 10.1016/j.plaphy.2025.109912. Epub 2025 Apr 10. Plant Physiol Biochem. 2025. PMID: 40239262
-
Identification of a new soybean kunitz trypsin inhibitor mutation and its effect on bowman-birk protease inhibitor content in soybean seed.J Agric Food Chem. 2015 Feb 11;63(5):1352-9. doi: 10.1021/jf505220p. Epub 2015 Jan 29. J Agric Food Chem. 2015. PMID: 25608918
Cited by
-
Enzymatic and Algebraic Methodology to Determine the Contents of Kunitz and Bowman-Birk Inhibitors and Their Contributions to Total Trypsin or Chymotrypsin Inhibition in Soybeans.J Agric Food Chem. 2024 May 22;72(20):11782-11793. doi: 10.1021/acs.jafc.3c06389. Epub 2024 May 8. J Agric Food Chem. 2024. PMID: 38717295 Free PMC article.
-
Long-read sequencing reveals novel structural variation markers for key agronomic and quality traits of food-grade soybean.Front Plant Sci. 2025 Apr 8;16:1557748. doi: 10.3389/fpls.2025.1557748. eCollection 2025. Front Plant Sci. 2025. PMID: 40265112 Free PMC article.
-
The Potential of CRISPR/Cas9 to Circumvent the Risk Factor Neurotoxin β-N-oxalyl-L-α, β-diaminopropionic acid Limiting Wide Acceptance of the Underutilized Grass Pea (Lathyrus sativus L.).Curr Issues Mol Biol. 2024 Sep 21;46(9):10570-10589. doi: 10.3390/cimb46090626. Curr Issues Mol Biol. 2024. PMID: 39329978 Free PMC article. Review.
-
Validation of Molecular Markers for Low Kunitz Trypsin Inhibitor Content in European Soybean (Glycine max L. Merr.) Germplasm.Genes (Basel). 2024 Aug 5;15(8):1028. doi: 10.3390/genes15081028. Genes (Basel). 2024. PMID: 39202388 Free PMC article.
-
Seed-Specific Silencing of Abundantly Expressed Soybean Bowman-Birk Protease Inhibitor Genes by RNAi Lowers Trypsin and Chymotrypsin Inhibitor Activities and Enhances Protein Digestibility.Int J Mol Sci. 2025 Jul 19;26(14):6943. doi: 10.3390/ijms26146943. Int J Mol Sci. 2025. PMID: 40725189 Free PMC article.
References
LinkOut - more resources
Full Text Sources